
Figure 4. Summary of growth cone-driven actin cytoskeletal changes in cells of the oligodendrocyte (OLG) lineage.
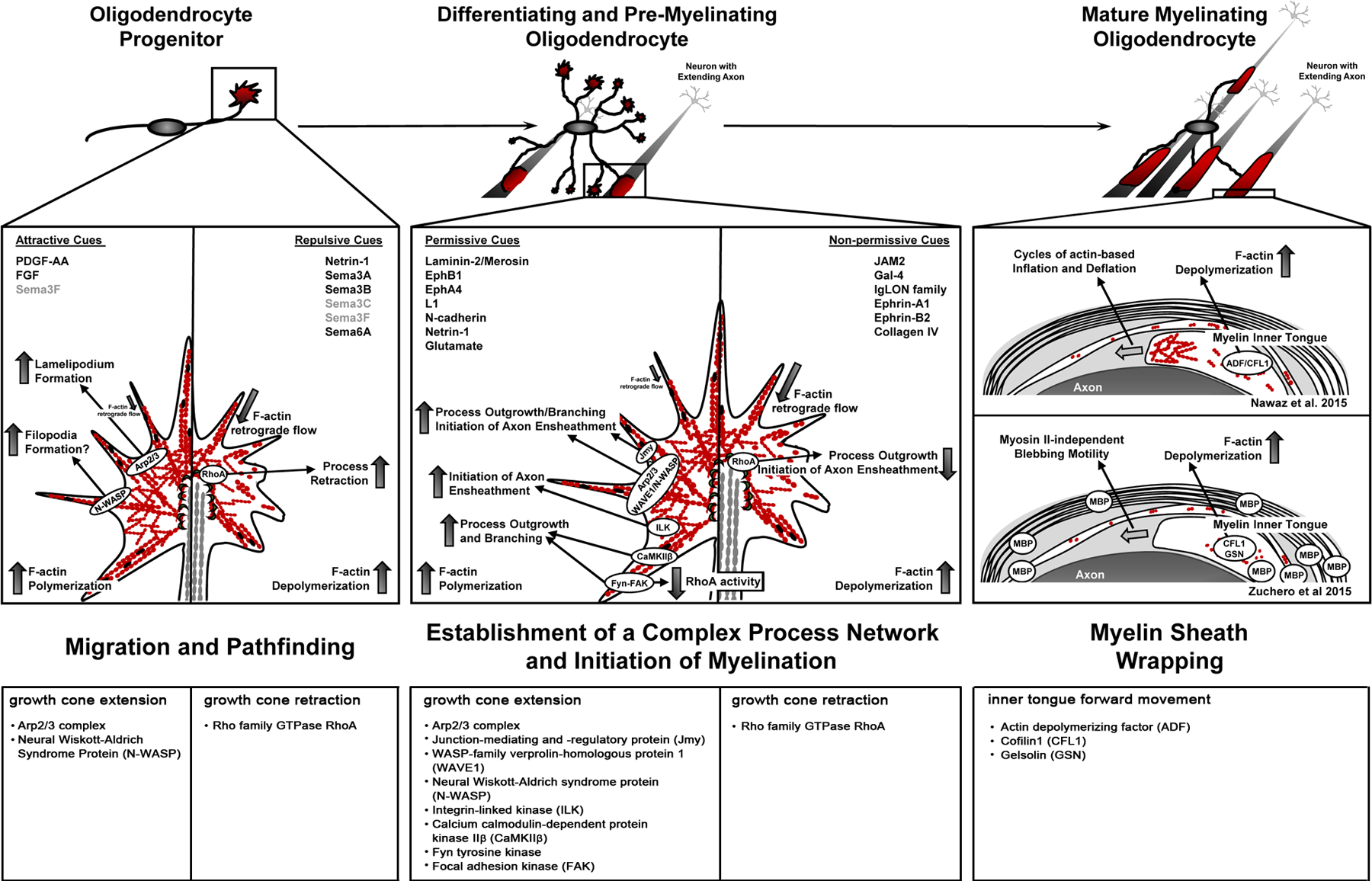
Cells of the OLG lineage undergo extensive changes in morphology when differentiating from a bipolar migratory OLG progenitor cell (OPC) first into a differentiating and pre-myelinating OLG that extends a large process network in search of axonal segments to be myelinated, and then into a mature OLG that generates and maintains a fully functional myelin sheath. At the progenitor stage (left), highly dynamic actin cytoskeletal mechanisms regulate growth cone behavior and, thereby, guide OPC migration along trajectories within the CNS in response to both attractive and repulsive extracellular cues. These motile behaviors are characterized by process extension events requiring F-actin polymerization, process retraction events driven by F-actin depolymerization, and process turning events that are mediated by an asymmetrical distribution of F-actin. Actin-binding proteins and key intracellular mediators implicated in OPC growth cone extension and retraction are depicted in the growth cone model (top) and listed in the bottom table. Differentiation into the differentiating OLG (middle) is associated with extensive process outgrowth and branching. In this process, F-actin polymerization and the actin regulatory proteins Arp2/3, Fyn, and focal adhesion kinase (FAK) play key roles in orchestrating actin cytoskeletal dynamics and motile behaviors (see growth cone model (top) and bottom table). Additional actin regulatory proteins, i.e. integrin-linked kinase (ILK), calcium calmodulin-dependent protein kinase IIβ (CaMKIIβ), and junction-mediating and -regulatory protein (Jmy), have been implicated in this process. However, the interactions, physical and/or functional, between all the potential players involved are currently not well understood. At the pre-myelinating stages, OLG processes respond to extracellular cues, which can be permissive as well as non-permissive, in an individual fashion. Growth cone responses at this stage are thought to be similar to the ones observed at the progenitor stage, and they are the determining factors regulating the initiation of myelination. The transition to the myelinating stage is associated with the loss of process extension events, as myelin wrapping is mediated largely by F-actin disassembly. F-actin severing proteins shown to be involved in this process are depicted in the model (top) and listed in the bottom table. Despite the consensus of F-actin disassembly as the major driver, there is still some controversy as to whether the force driving the myelin’s inner tongue is generated by cycles of actin-based inflation and deflation or by MBP-dependent, myosin II-independent blebbing motility.