

Abstract
Early caregiving experiences play a central role in shaping corticolimbic development and emotional learning and regulation. Given dynamic changes in corticolimbic maturation, the effects of caregiving experiences are likely to depend on the developmental timing of exposure. Cross-species evidence has identified timing-related differences in the effects of caregiving adversity. However, the extent to which developmental differences in associations between caregiving adversity and corticolimbic circuitry align with a sensitive period model has remained unclear. Converging evidence from studies of caregiver deprivation points to a sensitive period for caregiving influences on corticolimbic circuitry and emotional development during infancy. By contrast, differential associations between maltreatment and corticolimbic circuitry at specific ages in childhood and adolescence may reflect experience-dependent mechanisms of plasticity. Delineating sensitive periods of development and the precise experience-related mechanisms by which caregiving experiences influence corticolimbic development is essential for refining conceptual models and understanding risk and resilience following early adversity.
Keywords: caregiving, adversity, stress, brain development, corticolimbic circuitry, childhood, adolescence
Early experiences have profound and lasting effects on the developing brain and emotional behavior. Caregiving is one of the strongest species-expected inputs for altricial species early in life, and stable caregiving is critical to emotional well-being [1]. From normative variation in caregiving behaviors to severe caregiving adversity, the early environment actively shapes learning and behavioral repertoires in the affective domain for years to come. A rich cross-species literature has begun to identify the effects of caregiving experiences on corticolimbic circuitry and emotional behavior. Given marked changes in species-expected inputs and neuroplasticity across development, caregiving experiences are likely to differentially shape behavior during infancy, childhood, and adolescence [2–4]. However, much remains unknown about precise experience-related mechanisms and whether developmental differences in the effects of caregiving experiences reflect sensitive periods [5]. Identifying specific timing-related effects of caregiving experiences and differentiating between experience-expectant and experience-dependent mechanisms is key to advancing conceptual models of caregiving environments and corticolimbic development.
Sensitive Periods of Development
During a sensitive period of heightened neuroplasticity, a specific environmental input has a particularly strong influence on a specific brain circuit, and plasticity is limited following this window [6,7]. Importantly, sensitive periods are characterized by experience-expectant learning and are thought to reflect neural preparation to encode species-expected environmental stimuli [8]. Recent years have witnessed transformative discoveries of the molecular triggers (e.g., excitatory-inhibitory balance) and brakes (e.g., perineuronal nets, myelin) that control the onset and closure of sensitive periods, as well as the insight that sensitive period processes are themselves malleable [7]. Unlike experience-expectant plasticity, which tends to occur early in development, experience-dependent plasticity occurs in response to individual experiences (which are not necessarily species-expected) and facilitates learning throughout development [8]. The current review aims to apply a critical lens to existing research on caregiving effects on corticolimbic development to begin to delineate which developmental differences in caregiving influences may align with a sensitive period model and experience-expectant versus experience-dependent plasticity.
Caregiving and Corticolimbic Development
Decades of research have demonstrated the robust links between caregiving and offspring emotional behavior [9], with a growing literature focused on neurobiological mechanisms. Cross-species evidence has demonstrated that early caregiving experiences have particularly strong effects on corticolimbic circuity involved in learning about salient aspects of the environment and regulating emotion. Connections between regions such as the medial prefrontal cortex (mPFC), amygdala, and hippocampus, which play a key role in regulating emotion and guiding biologically-relevant learning [10], may be especially impacted by adversity due to their dense innervation with glucocorticoid receptors and the developmental timing of circuit maturation [11]. Whereas prefrontal regions and their connections with limbic structures undergo protracted development, the amygdala matures relatively earlier and may be particularly sensitive to the early social environment [12]. Though these connections undergo marked changes across development [13,14], functional connectivity at rest is already evident between the amygdala and regions such as ventromedial prefrontal cortex (vmPFC) in infancy [15]. Moreover, amygdala-mPFC functional connectivity at rest among newborns is associated with negative affect at six months of age [16] and with behavioral inhibition at two years of age [17]. Environmental influences on corticolimbic circuitry in early life may play an active role in shaping longer-term neural and behavioral phenotypes. For example, neural co-activations induced via experiences with caregivers may “entrain” the system during a highly plastic time in ways that shape intrinsic corticolimbic architecture and affective behaviors [18–20]. Moreover, the early sensitivity of the amygdala to environmental inputs may directly guide mPFC function and connectivity [20] and influence later-developing aspects of broader cortico-subcortical circuitry through developmental cascades [21].
Much of the research linking caregiving with emotional development comes from studies of severe caregiving adversity. Alterations of the HPA axis [22] and corticolimbic circuitry [23,24] appear to underlie effects of caregiving adversity on emotional learning [25] and regulation [26] and likely contribute to increased risk for mental health disorders. However, increasingly evidence has also emerged linking normative variation in caregiving behavior with corticolimbic structure and function [27]. For example, during childhood, caregiver sensitivity is associated with amygdala volume and microstructure of the amygdala and hippocampus [28], and negative caregiving behavior is associated with amygdala activation and functional connectivity between the amygdala and superior parietal lobule [29]. In addition, caregiver control experienced during childhood is associated with amygdala activation and structural integrity of the uncinate fasciculus during young adulthood [30]. These studies further underscore the importance of caregiving in healthy brain development.
Developmental Differences in the Effects of Caregiving on Corticolimbic Circuitry
Non-human animal work that allows for manipulating the timing of exposure shows that the effects of stress differ as a function of developmental timing [3]. Manipulating stress exposure is challenging in humans; however, naturalistic human studies of unfortunate events (e.g., institutionalized care) provide strong evidence for the importance of the timing of adversity. With its unique study design, the Bucharest Early Intervention Project has highlighted a potential sensitive period related to socioemotional development during the first two years of life. That is, youth who were exposed to caregiver deprivation via institutionalized care show more secure attachment, more normative stress responses, and more normative neurodevelopmental trajectories following placement into a foster care intervention prior to 24 months of age, relative to peers who were placed later [31]. These findings highlight infancy as a particularly important time for caregiving influences, as well as the potential for the identification of sensitive periods to inform interventions.
Consistent evidence has shown that the absence of stable, nurturing caregiving in the postnatal and infancy period disrupts corticolimbic development. Across species, early caregiver deprivation is associated with altered connectivity between the amygdala and mPFC in mice [32], rats [33], non-human primates [23], and humans [24]. It is possible that these findings reflect a sensitive period driven by experience-expectant mechanisms. Consistent with criteria for a sensitive period [7,34], infancy is a time of rapid and marked change in corticolimbic circuitry [35], and it is biologically plausible that this period is characterized by heightened neuroplasticity. There is also some specificity to the nature of the experience, the neural circuit affected, and the timing of the window during which caregiver deprivation has particularly strong effects [3]. However, evaluating sensitive period phenomena in human development is especially challenging, and additional research will be necessary to more rigorously assess all relevant criteria [5,34]. In particular, it is rare that longitudinal data are available to test whether effects of caregiver deprivation on corticolimbic circuitry persist into adulthood.
Although substantial evidence suggests that caregiving adversity has the strongest effects when experienced earlier in life [3,22,31], an alternative account suggests that risk may be highest when adversity occurs during specific windows that could occur later in childhood or adolescence. Studies examining variation in the timing of maltreatment have at times pointed to specific ages of exposure during childhood or adolescence at which effects on corticolimbic structure or function in adulthood are pronounced [36–39]. These studies highlight the complexity of interactions between developmental timing with the type of adversity exposure, sex, and regional specificity in the brain. For example, exposure to maltreatment between ages 10 and 11 is specifically related to amygdala volume in adulthood, relative to exposure at other ages during development [37], whereas sexual abuse at ages 3–5 and 11–13 is uniquely associated with hippocampal volume in adulthood [36]. Among men, hippocampal volume in adulthood is associated with neglect, but not abuse, prior to age 7. By contrast, hippocampal volume in women is associated with abuse, but not neglect, at ages 10–11 and 15–16 years [38]. However, it is not clear whether these developmental differences align with a sensitive period process, and experience-dependent mechanisms may better explain such age-related effects. As one example, maltreatment is unlikely to be a plausible type of species-expected stimuli at a specific developmental time [5]. Moreover, while these findings in adulthood may suggest non-linear peaks in risk throughout development, future studies during childhood and adolescence will be important for understanding more proximal corticolimbic changes that may unfold across development.
Yet another way in which caregiving adversity may alter corticolimbic development is by altering sensitive period processes themselves. In rodents, hippocampal and amygdala development, as well as some forms of emotional learning, are accelerated following early adversity [40–42]. In humans, evidence suggests that the timing of structural and functional corticolimbic development may also be accelerated following adversity [24,43,44]. As one example, while viewing fearful faces, children who experienced caregiver deprivation exhibit more mature patterns of functional connectivity between the vmPFC and amygdala (i.e., negative task-based connectivity), which resemble those of adolescents and adults [24]. Across species these effects have been mediated by corticosterone levels (cortisol in humans) [24], suggesting that early caregiving adversity may prematurely stimulate the HPA axis in a way that contributes to precocious corticolimbic and emotional development.
Such accelerated development may represent an ontogenetic adaptation in the context of an early harsh environment [45,46]. Consistent with this idea, children exposed to caregiver deprivation who show the more mature phenotype of vmPFC-amygdala connectivity also display lower separation anxiety [24]. These findings are in line with evidence that stronger inverse amygdala-mPFC functional connectivity is associated with lower internalizing symptoms among youth exposed to early family adversity [47] and that greater prefrontal control (specifically, superior frontal gyrus and dorsal anterior cingulate cortex) of amygdala reactivity during emotion regulation is associated with lower depressive symptoms following child maltreatment [48]. Moreover, recent work shows that the more mature pattern of vmPFC-amygdala connectivity is also associated with slower telomere shortening and pubertal tempo [*49], which may further suggest protective effects in the context of evidence demonstrating accelerated cellular aging following early-life adversity. However, there are likely to be long-term consequences of accelerated development. Such precocious maturation may signal a shift or premature termination of a sensitive period of caregiving influences, which could be associated with reduced plasticity. Future research will be important for understanding longer-term effects, testing whether neural findings are specific to corticolimbic circuitry, and for further examining how developmental patterns of acceleration converge or diverge across different domains such as puberty, cellular aging, and neurodevelopment.
Early Caregiving Influences on Emotional Learning and Regulation
Caregivers modulate offspring behavior in a number of domains related to affective behavior, including influencing what offspring learn and how they perceive the world around them. Encoding stable, reliable caregiver cues that are associated with safety during an early sensitive period (e.g., infancy) may be essential to the roles that caregivers play in modulating emotional learning and regulation later in development. Young offspring show a preference for cues related to their caregiver, even when those cues are inherently aversive. For example, rodent pups show approach behaviors toward an odor paired with a shock during a period when maternal presence maintains low levels of corticosterone and blocks amygdala plasticity [50]. Paralleling these findings in rodents, recent evidence demonstrates similar caregiver-related learning in humans. Specifically, young children were more likely to approach conditioned stimuli that were acquired in their caregiver’s presence and to avoid stimuli acquired in the caregiver’s absence [*51]. This attraction to caregiver-related cues and absence of avoidance learning is thought to facilitate attachment early in life and ensure that the offspring stays close to their caregiver [52]. Critically, these effects depend on developmental stage. During a window from postnatal day (P)10 to P15, rodent pup behavior depends on maternal presence, such that pups continue to show approach behaviors if the mother is present. However, corticosterone and amygdala activation increase if the mother is absent, instantiating threat learning and avoidance behaviors [50], potentially to facilitate survival when offspring engage in exploration independently.
Consistent with the effects of caregiver presence on corticosterone levels and amygdala plasticity in rodents, caregivers buffer stress physiology and HPA axis reactivity in infant macaques [53] and in humans [54]. Suggesting a potentially related mechanism by which caregivers modulate affective behavior early in life, during childhood, caregivers suppress amygdala reactivity and phasically induce a pattern of amygdala-mPFC functional connectivity that may be more strongly regulatory [55]. Paralleling this modulation of amygdala-mPFC circuitry, children also show enhanced regulatory behavior in an affective context in the presence of their mother compared with a stranger. The effects of caregivers on cortisol reactivity [54] as well as on regulatory behavior and amygdala-mPFC circuitry [55] are specific to childhood, and not adolescence. These findings suggest that caregivers may serve an external regulatory function while corticolimbic circuitry is still developing. With time and as this circuitry matures, reliance on external regulation may decrease as regulatory abilities become internalized to facilitate independent emotion regulation, and other major attachment figures such as close peers or romantic partners may take on an increasing role in social buffering [46,56] (Figure 1). In humans, caregiver presence has also been shown to increase discrimination between threat and safety cues during childhood, but not adolescence [57]. In these ways, caregivers play a central role in shaping what children learn about their environment and in regulating their behavior, particularly early in life. However, instead of caregiver effects on emotional learning and regulation during childhood reflecting a sensitive period itself, it may be that encoding reliable cues related to the support of caregivers earlier in life allows for those cues to exert these unique effects during childhood.
Figure 1. Caregiver influences on corticolimbic circuitry related to emotion regulation across development.
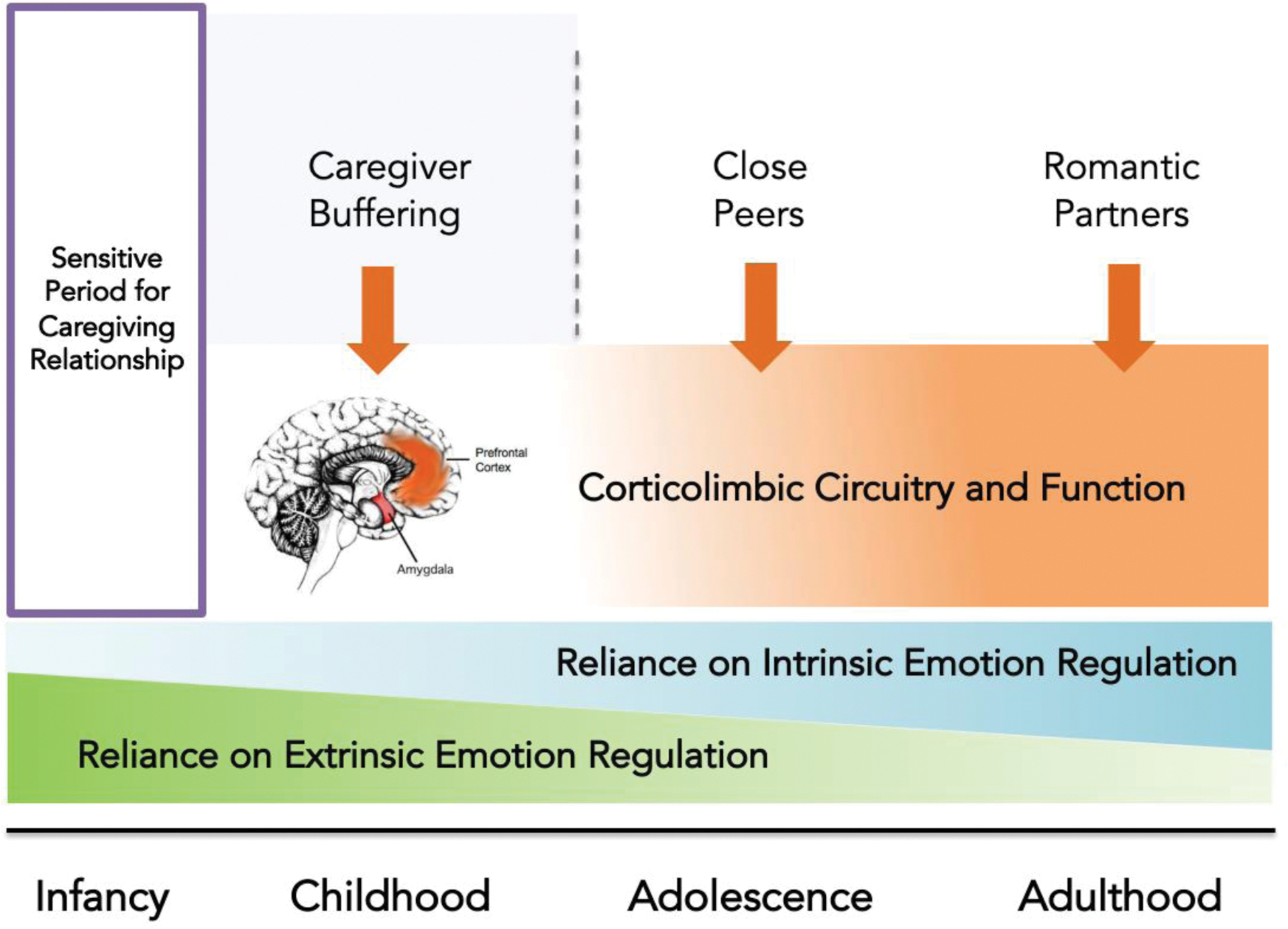
Cross-species evidence suggests a sensitive period during infancy through which caregiving has particularly strong influences on corticolimbic development (including mPFC-amygdala connectivity) and longer-term emotional behavior. Caregivers regulate amygdala function during infancy (demonstrated in rodents, hypothesized in humans) and childhood (demonstrated in humans), and not in adolescence. The shift away from reliance on caregivers for extrinsic regulation may be accompanied by increased capacity for intrinsic emotion regulation around the transition to adolescence. Social buffering continues across the lifespan, with different primary attachment figures potentially serving a regulatory function at distinct developmental stages. Due to heightened plasticity to caregiving influences early in life, severe disruptions in caregiving during infancy may interfere with learning reliable, safe caregiver cues in a way that interferes with normative caregiver shaping of emotional learning and regulation later in development. Adapted with permission from Gee, 2016.
Consistent with this idea, increasing cross-species evidence suggests that early caregiving adversity disrupts the ways in which caregivers guide learning and buffer emotional reactivity in later stages of development. In rodents, infant maltreatment is associated with reduced effects of maternal presence on threat learning during infancy [58,*59]. Interestingly, maltreatment during infancy has differential effects on maternal buffering during infancy versus adolescence. Whereas maltreatment completely disrupts maternal buffering at PN18, maternal buffering is present but weaker at PN28 [*59]. In non-human primates, infant maltreatment is associated with weaker maternal buffering of stress-induced cortisol increases [60]. In humans, caregiver deprivation early in life interferes with caregiver buffering of amygdala reactivity during childhood [**61]. However, effects are heterogeneous, such that approximately 40% of youth who experienced caregiver deprivation exhibit caregiver buffering, and those youth also experience steeper declines in separation anxiety over a period of three years. Thus, the ability to experience caregiver buffering of amygdala reactivity may enhance resilience within this group at elevated risk for anxiety. Taken together, these findings suggest that while caregiver buffering itself may not be consistent with a sensitive period phenomenon, caregiving adversity during a sensitive period in infancy may disrupt the encoding of stable, safe caregiver cues that are likely to be important for caregiver influences on emotional learning and regulation later in development.
Future Directions
Despite a growing literature on developmental differences in the effects of caregiving on corticolimbic development, the experience-related mechanisms underlying these influences remain largely unknown. Ongoing research that evaluates which developmental differences are consistent with the criteria for a sensitive period will be essential to advancing conceptual models and understanding the mechanisms by which early caregiving experiences become biologically embedded to shape emotional development. It is rare that distinct models of experience-related mechanisms have been directly compared, and rigorously testing a sensitive period model in humans presents various challenges [5,62], including the complex and multifaceted nature of caregiving experiences, the protracted time needed to assess effects on mature function, and the inability to test for molecular regulators and direct markers of plasticity in humans. In addition, it is important to acknowledge that experience-expectant and experience-dependent learning are unlikely to be completely independent processes [8], and experience-related learning is unlikely to reflect a single process or model. In this regard, cross-species research will continue to be essential to testing hypotheses about sensitive periods [63], and bridging between formal modeling and empirical studies may offer powerful insights [62]. Further, study designs will need to incorporate precise measurement of timing of exposures and extend longitudinal follow-up to enhance the ability to test sensitive period models in human neurodevelopment.
Lastly, research on developmental differences in caregiving effects has largely focused on timing-related factors in isolation. An important area for future research will be examining how timing-related factors (e.g., age of exposure, chronicity, duration) interact with key experiential dimensions of adversity, such as the extent to which adversity is characterized by threat versus deprivation [64], predictability [65], controllability, and/or caregiver involvement [for review, see **66]. As one example, independent of severity, adversity perpetrated by a caregiver or adversity that involves dyadic caregiver/child exposure may have stronger or differential effects on corticolimbic development than adversity that does not involve a caregiver [67]. Further delineating how specific features of adversity differentially impact outcomes, and how those effects differ by developmental stage, could inform efforts to optimize risk identification based on developmental stage or the nature of adversity exposure.
Conclusions
Cross-species findings demonstrate that early caregiving experiences play a central role in shaping the development of learning and regulation in the affective domain. Adverse caregiving can alter corticolimbic development and normative processes such as caregiver buffering of amygdala reactivity, with lasting implications for emotional behavior and mental health. While increasing evidence demonstrates that caregiving influences depend on the timing of experiences, it is unclear under what circumstances earlier adversity is more consequential or whether there are windows of development throughout childhood and adolescence when caregiving adversity has the strongest effects. Moreover, much remains unknown about the experience-related mechanisms of plasticity that link early caregiving inputs with affective outcomes. Caregiver deprivation experienced during infancy has persistent effects on corticolimbic development and later caregiver buffering, which may reflect an early sensitive period for attachment and learning stable caregiver cues. Future research will be essential for testing whether developmental differences in caregiving influences may reflect experience-expectant or experience-dependent mechanisms, or influences on sensitive period timing itself. Refining conceptual models based on such knowledge has important implications for promoting resilience following early adversity and could be leveraged to enhance risk identification or tailor interventions based on factors such as developmental stage.
Highlights.
Caregiving experiences influence corticolimbic circuitry and emotional development
Effects of adverse caregiving depend on the timing of exposure
Infancy may represent a sensitive period (experience-expectant learning) for caregiver deprivation
Experience-dependent mechanisms may underlie effects of maltreatment later in development
Translating such knowledge may enhance risk identification and interventions
Acknowledgements
This work was supported by the National Institutes of Health (NIH) Director’s Early Independence Award (DP5OD021370), Brain & Behavior Research Foundation (National Alliance for Research on Schizophrenia and Depression; NARSAD) Young Investigator Award, Jacobs Foundation Early Career Research Fellowship, and the Society for Clinical Child and Adolescent Psychology (Division 53 of the American Psychological Association) Richard “Dick” Abidin Early Career Award and Grant.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
The author has no conflicts of interest to disclose.
References
- 1.Tottenham N: Human amygdala development in the absence of species-expected caregiving. Developmental Psychobiology 2012, 54:598–611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gee DG, Casey BJ: The Impact of Developmental Timing for Stress and Recovery. Neurobiol Stress 2015, 1:184–194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lupien SJ, McEwen BS, Gunnar MR, Heim C: Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat Rev Neurosci 2009, 10:434–445. [DOI] [PubMed] [Google Scholar]
- 4.Teicher MH, Samson JA, Anderson CM, Ohashi K: The effects of childhood maltreatment on brain structure, function and connectivity. Nature Reviews Neuroscience 2016, 17:652–666. [DOI] [PubMed] [Google Scholar]
- 5.Gabard-Durnam LJ, McLaughlin KA: Do Sensitive Periods Exist for Exposure to Adversity? Biol Psychiatry 2019, 85:789–791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Knudsen EI: Sensitive periods in the development of the brain and behavior. J Cogn Neurosci 2004, 16:1412–1425. [DOI] [PubMed] [Google Scholar]
- 7.Werker JF, Hensch TK: Critical Periods in Speech Perception: New Directions. Annual Review of Psychology 2015, 66:173–196. [DOI] [PubMed] [Google Scholar]
- 8.Greenough WT, Black JE, Wallace CS: Experience and Brain Development. Child Development 1987, [PubMed] [Google Scholar]
- 9.Hofer MA: Hidden Regulators in Attachment, Separation, and Loss. Monographs of the Society for Research in Child Development 1994, 59:192–207. [PubMed] [Google Scholar]
- 10.Kovner R, Oler JA, Kalin NH: Cortico-Limbic Interactions Mediate Adaptive and Maladaptive Responses Relevant to Psychopathology. AJP 2019, 176:987–999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tottenham N, Sheridan M: A review of adversity, the amygdala and the hippocampus: a consideration of developmental timing. Front Hum Neurosci 2009, 3:68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Raineki C, Opendak M, Sarro E, Showler A, Bui K, McEwen BS, Wilson DA, Sullivan RM: During infant maltreatment, stress targets hippocampus, but stress with mother present targets amygdala and social behavior. Proc Natl Acad Sci USA 2019, 116:22821–22832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gabard-Durnam LJ, Flannery J, Goff B, Gee DG, Humphreys KL, Telzer E, Hare T, Tottenham N: The development of human amygdala functional connectivity at rest from 4 to 23years: A cross-sectional study. Neuroimage 2014, doi: 10.1016/j.neuroimage.2014.03.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gee DG, Humphreys KL, Flannery J, Goff B, Telzer EH, Shapiro M, Hare TA, Bookheimer SY, Tottenham N: A developmental shift from positive to negative connectivity in human amygdala-prefrontal circuitry. J Neurosci 2013, 33:4584–4593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gabard-Durnam LJ, O’Muircheartaigh J, Dirks H, Dean DC, Tottenham N, Deoni S: Human amygdala functional network development: A cross-sectional study from 3 months to 5 years of age. Dev Cogn Neurosci 2018, 34:63–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Graham AM, Buss C, Rasmussen JM, Rudolph MD, Demeter DV, Gilmore JH, Styner M, Entringer S, Wadhwa PD, Fair DA: Implications of newborn amygdala connectivity for fear and cognitive development at 6-months-of-age. Developmental Cognitive Neuroscience 2016, 18:12–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rogers CE, Sylvester CM, Mintz C, Kenley JK, Shimony JS, Barch DM, Smyser CD: Neonatal Amygdala Functional Connectivity at Rest in Healthy and Preterm Infants and Early Internalizing Symptoms. Journal of the American Academy of Child & Adolescent Psychiatry 2017, 56:157–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Callaghan BL, Tottenham N: The Neuro-Environmental Loop of Plasticity: A Cross-Species Analysis of Parental Effects on Emotion Circuitry Development Following Typical and Adverse Caregiving. Neuropsychopharmacology 2016, 41:163–176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gabard-Durnam LJ, Gee DG, Goff B, Flannery J, Telzer E, Humphreys KL, Lumian DS, Fareri DS, Caldera C, Tottenham N: Stimulus-Elicited Connectivity Influences Resting-State Connectivity Years Later in Human Development: A Prospective Study. J Neurosci 2016, 36:4771–4784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Tottenham N, Gabard-Durnam L: The developing amygdala: a student of the world and a teacher of the cortex. Current Opinion in Psychology 2017, 17:55–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Casey BJ, Heller AS, Gee DG, Cohen AO: Development of the emotional brain. Neurosci Lett 2019, 693:29–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gunnar MR, Frenn K, Wewerka SS, Ryzin MJV: Moderate versus severe early life stress: Associations with stress reactivity and regulation in 10–12-year-old children. Psychoneuroendocrinology 2009, 34:62–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Howell BR, Ahn M, Shi Y, Godfrey JR, Hu X, Zhu H, Styner M, Sanchez MM: Disentangling the effects of early caregiving experience and heritable factors on brain white matter development in rhesus monkeys. Neuroimage 2019, 197:625–642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gee DG, Gabard-Durnam LJ, Flannery J, Goff B, Humphreys KL, Telzer EH, Hare TA, Bookheimer SY, Tottenham N: Early developmental emergence of human amygdala-prefrontal connectivity after maternal deprivation. Proc Natl Acad Sci USA 2013, 110:15638–15643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McLaughlin KA, Sheridan MA, Gold AL, Duys A, Lambert HK, Peverill M, Heleniak C, Shechner T, Wojcieszak Z, Pine DS: Maltreatment Exposure, Brain Structure, and Fear Conditioning in Children and Adolescents. Neuropsychopharmacology 2016, 41:1956–1964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Heleniak C, Jenness JL, Stoep AV, McCauley E, McLaughlin KA: Childhood Maltreatment Exposure and Disruptions in Emotion Regulation: A Transdiagnostic Pathway to Adolescent Internalizing and Externalizing Psychopathology. Cognit Ther Res 2016, 40:394–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Farber MJ, Gee DG, Hariri AR: Normative range parenting and the developing brain: A scoping review and recommendations for future research. Eur J Neurosci 2020, doi: 10.1111/ejn.15003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lee A, Poh JS, Wen DJ, Tan HM, Chong Y-S, Tan KH, Gluckman PD, Fortier MV, Rifkin-Graboi A, Qiu A: Maternal care in infancy and the course of limbic development. Developmental Cognitive Neuroscience 2019, 40:100714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pozzi E, Simmons JG, Bousman CA, Vijayakumar N, Bray KO, Dandash O, Richmond S, Schwartz O, Seal M, Sheeber L, et al. : The Influence of Maternal Parenting Style on the Neural Correlates of Emotion Processing in Children. Journal of the American Academy of Child & Adolescent Psychiatry 2019, doi: 10.1016/j.jaac.2019.01.018. [DOI] [PubMed] [Google Scholar]
- 30.Farber MJ, Kim MJ, Knodt AR, Hariri AR: Maternal overprotection in childhood is associated with amygdala reactivity and structural connectivity in adulthood. Developmental Cognitive Neuroscience 2019, 40:100711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.McLaughlin KA, Sheridan MA, Tibu F, Fox NA, Zeanah CH, Nelson CA: Causal effects of the early caregiving environment on development of stress response systems in children. PNAS 2015, doi: 10.1073/pnas.1423363112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Johnson FK, Delpech J-C, Thompson GJ, Wei L, Hao J, Herman P, Hyder F, Kaffman A: Amygdala hyper-connectivity in a mouse model of unpredictable early life stress. Translational Psychiatry 2018, 8:49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yan C-G, Rincón-Cortés M, Raineki C, Sarro E, Colcombe S, Guilfoyle DN, Yang Z, Gerum S, Biswal BB, Milham MP, et al. : Aberrant development of intrinsic brain activity in a rat model of caregiver maltreatment of offspring. Transl Psychiatry 2017, 7:e1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Takesian AE, Hensch TK: Balancing plasticity/stability across brain development. Prog Brain Res 2013, 207:3–34. [DOI] [PubMed] [Google Scholar]
- 35.Payne C, Machado CJ, Bliwise NG, Bachevalier J: Maturation of the Hippocampal Formation and Amygdala in Macaca mulatta: A Volumetric Magnetic Resonance Imaging Study. Hippocampus 2010, 20:922–935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Andersen SL, Tomada A, Vincow ES, Valente E, Polcari A, Teicher MH: Preliminary evidence for sensitive periods in the effect of childhood sexual abuse on regional brain development. J Neuropsychiatry Clin Neurosci 2008, 20:292–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pechtel P, Lyons-Ruth K, Anderson CM, Teicher MH: Sensitive periods of amygdala development: the role of maltreatment in preadolescence. Neuroimage 2014, 97:236–244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Teicher MH, Anderson CM, Ohashi K, Khan A, McGreenery CE, Bolger EA, Rohan ML, Vitaliano GD: Differential effects of childhood neglect and abuse during sensitive exposure periods on male and female hippocampus. Neuroimage 2018, 169:443–452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *39.Zhu J, Lowen SB, Anderson CM, Ohashi K, Khan A, Teicher MH: Association of Prepubertal and Postpubertal Exposure to Childhood Maltreatment With Adult Amygdala Function. JAMA Psychiatry 2019, doi: 10.1001/jamapsychiatry.2019.0931. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study highlights the importance of developmental timing of adversity exposure, as well as how timing may interact with other key features of exposure such as the type of adversity. Specifically, the authors find that physical abuse experienced between ages 3–6 is associated with blunted amygdala reactivity in adulthood, whereas peer emotional abuse at ages 13 and 15 is associated with heightened amygdala reactivity in adulthood.
- 40.Bath KG, Manzano-Nieves G, Goodwill H: Early life stress accelerates behavioral and neural maturation of the hippocampus in male mice. Horm Behav 2016, 82:64–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Callaghan BL, Richardson R: Maternal separation results in early emergence of adult-like fear and extinction learning in infant rats. Behav Neurosci 2011, 125:20–28. [DOI] [PubMed] [Google Scholar]
- 42.Manzano Nieves G, Bravo M, Baskoylu S, Bath KG: Early life adversity decreases pre-adolescent fear expression by accelerating amygdala PV cell development. eLife 2020, 9:e55263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Colich NL, Williams ES, Ho TC, King LS, Humphreys KL, Price AN, Ordaz SJ, Gotlib IH: The Association Between Early Life Stress and Prefrontal Cortex Activation During Implicit Emotion Regulation is Moderated by Sex in Early Adolescence. Dev Psychopathol 2017, 29:1851–1864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Silvers JA, Lumian DS, Gabard-Durnam L, Gee DG, Goff B, Fareri DS, Caldera C, Flannery J, Telzer EH, Humphreys KL, et al. : Previous Institutionalization Is Followed by Broader Amygdala-Hippocampal-PFC Network Connectivity during Aversive Learning in Human Development. J Neurosci 2016, 36:6420–6430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Callaghan BL, Tottenham N: The Stress Acceleration Hypothesis: effects of early-life adversity on emotion circuits and behavior. Current Opinion in Behavioral Sciences 2016, 7:76–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gee DG: Sensitive Periods of Emotion Regulation: Influences of Parental Care on Frontoamygdala Circuitry and Plasticity. New Dir Child Adolesc Dev 2016, 2016:87–110. [DOI] [PubMed] [Google Scholar]
- 47.Herringa RJ, Burghy CA, Stodola DE, Fox ME, Davidson RJ, Essex MJ: Enhanced prefrontal-amygdala connectivity following childhood adversity as a protective mechanism against internalizing in adolescence. Biol Psychiatry Cogn Neurosci Neuroimaging 2016, 1:326–334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rodman AM, Jenness JL, Weissman DG, Pine DS, McLaughlin KA: Neurobiological markers of resilience to depression and anxiety following childhood maltreatment: The role of neural circuits supporting the cognitive control of emotion. Biological Psychiatry 2019, 0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *49.Miller JG, Ho TC, Humphreys KL, King LS, Foland-Ross LC, Colich NL, Ordaz SJ, Lin J, Gotlib IH: Early Life Stress, Frontoamygdala Connectivity, and Biological Aging in Adolescence: A Longitudinal Investigation. Cereb Cortex 2020, 30:4269–4280. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this longitudinal study, the authors find that greater cumulative severity of early adversity is associated with stronger negative coupling between the amygdala and ventromedial prefrontal cortex. These findings are consistent with prior cross-sectional research (Gee et al., 2013) and may indicate accelerated neurodevelopment. Further, this study examined cellular aging and pubertal tempo, and findings suggest that frontolimbic changes following adversity may reflect a different developmental process.
- 50.Moriceau S, Sullivan RM: Maternal presence serves as a switch between learning fear and attraction in infancy. Nat Neurosci 2006, 9:1004–1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *51.Tottenham N, Shapiro M, Flannery J, Caldera C, Sullivan RM: Parental presence switches avoidance to attraction learning in children. Nat Hum Behav 2019, doi: 10.1038/s41562-019-0656-9. [DOI] [PMC free article] [PubMed] [Google Scholar]; Consistent with evidence in rodents, this study demonstrates that affective learning is shaped by caregivers early in human development. Specifically, young children showed greater behavioral approach to a conditioned stimulus that was acquired in their caregiver’s presence and avoidance of a conditioned stimulus acquired in their caregiver’s absence.
- 52.Perry R, Sullivan RM: Neurobiology of attachment to an abusive caregiver: short-term benefits and long-term costs. Dev Psychobiol 2014, 56:1626–1634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sanchez MM: The impact of early adverse care on HPA axis development: nonhuman primate models. Horm Behav 2006, 50:623–631. [DOI] [PubMed] [Google Scholar]
- 54.Hostinar CE, Johnson AE, Gunnar MR: Parent support is less effective in buffering cortisol stress reactivity for adolescents compared to children. Dev Sci 2015, 18:281–297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Gee DG, Gabard-Durnam L, Telzer EH, Humphreys KL, Goff B, Shapiro M, Flannery J, Lumian DS, Fareri DS, Caldera C, et al. : Maternal buffering of human amygdala-prefrontal circuitry during childhood but not during adolescence. Psychol Sci 2014, 25:2067–2078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hornstein EA, Eisenberger NI: Unpacking the buffering effect of social support figures: Social support attenuates fear acquisition. PLoS One 2017, 12:e0175891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.van Rooij SJH, Cross D, Stevens JS, Vance LA, Kim YJ, Bradley B, Tottenham N, Jovanovic T: Maternal buffering of fear-potentiated startle in children and adolescents with trauma exposure. Soc Neurosci 2017, 12:22–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Opendak M, Robinson-Drummer P, Blomkvist A, Zanca RM, Wood K, Jacobs L, Chan S, Tan S, Woo J, Venkataraman G, et al. : Neurobiology of maternal regulation of infant fear: the role of mesolimbic dopamine and its disruption by maltreatment. Neuropsychopharmacology 2019, 44:1247–1257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *59.Robinson-Drummer PA, Opendak M, Blomkvist A, Chan S, Tan S, Delmer C, Wood K, Sloan A, Jacobs L, Fine E, et al. : Infant Trauma Alters Social Buffering of Threat Learning: Emerging Role of Prefrontal Cortex in Preadolescence. Front Behav Neurosci 2019, 13:132. [DOI] [PMC free article] [PubMed] [Google Scholar]; Building upon a rich cross-species literature on caregiver buffering, this study in rodents examined the effect of maltreatment on maternal buffering of threat learning. The authors find that maltreatment blocks maternal buffering of pups’ threat learning during infancy. The findings provide important insight into how the effects of early adversity differ depending on developmental stage. Specifically, the effect of maltreatment on buffering appears to differ during adolescence, such that maltreatment attenuates but does not block threat learning.
- 60.Sanchez MM, McCormack KM, Howell BR: Social Buffering of Stress Responses in Nonhuman Primates: Maternal Regulation of the Development of Emotional Regulatory Brain Circuits. Soc Neurosci 2015, 10:512–526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- **61.Callaghan BL, Gee DG, Gabard-Durnam L, Telzer EH, Humphreys KL, Goff B, Shapiro M, Flannery J, Lumian DS, Fareri DS, et al. : Decreased Amygdala Reactivity to Parent Cues Protects Against Anxiety Following Early Adversity: An Examination Across 3 Years. Biol Psychiatry Cogn Neurosci Neuroimaging 2019, 4:664–671. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study examined caregiver buffering of amygdala reactivity among children and adolescents previously exposed to parental deprivation. The authors find that severe caregiving-related adversity disrupts the normative process of caregiver buffering. However, a substantial proportion of youth exposed to parental deprivation do show caregiver buffering of amygdala reactivity, which is associated with lower risk of anxiety.
- 62.Frankenhuis WE, Walasek N: Modeling the Evolution of Sensitive Periods. Developmental Cognitive Neuroscience 2019, doi: 10.1016/j.dcn.2019.100715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Opendak M, Gould E, Sullivan R: Early life adversity during the infant sensitive period for attachment: Programming of behavioral neurobiology of threat processing and social behavior. Dev Cogn Neurosci 2017, 25:145–159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sheridan MA, McLaughlin KA: Dimensions of early experience and neural development: deprivation and threat. Trends Cogn Sci (Regul Ed) 2014, 18:580–585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Baram TZ, Solodkin A, Davis EP, Stern H, Obenaus A, Sandman CA, Small SL: Fragmentation and Unpredictability of Early-Life Experience in Mental Disorders. Am J Psychiatry 2012, 169:907–915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- **66.Cohodes EM, Kitt ER, Baskin-Sommers A, Gee DG: Influences of early-life stress on frontolimbic circuitry: Harnessing a dimensional approach to elucidate the effects of heterogeneity in stress exposure. Developmental Psychobiology 2020. [DOI] [PubMed] [Google Scholar]; This review examines heterogeneity in early adversity and proposes a framework by which key dimensions of early adversity may differentially impact the development of frontolimbic circuitry. The authors particularly focus on the developmental timing of adversity exposure and how timing may interact with other factors such as caregiver involvement to influence outcomes.
- 67.Cook A, Spinazzola J, Ford J, Lanktree C, Blaustein M, Cloitre M, DeRosa R, Hubbard R, Kagan R, Liautaud J, et al. : Complex Trauma in Children and Adolescents. Psychiatric Annals 2005, 35:390–398. [Google Scholar]