

SUMMARY
Conflicts between the replication and transcription machineries have profound effects on chromosome duplication, genome organization, and evolution across species. Head-on conflicts (lagging-strand genes) are significantly more detrimental than codirectional conflicts (leading-strand genes). The fundamental reason for this difference is unknown. Here, we report that topological stress significantly contributes to this difference. We find that head-on, but not codirectional, conflict resolution requires the relaxation of positive supercoils by the type II topoisomerases DNA gyrase and Topo IV, at least in the Gram-positive model bacterium Bacillus subtilis. Interestingly, our data suggest that after positive supercoil resolution, gyrase introduces excessive negative supercoils at head-on conflict regions, driving pervasive R-loop formation. Altogether, our results reveal a fundamental mechanistic difference between the two types of encounters, addressing a long-standing question in the field of replication-transcription conflicts.
Graphical Abstract
In brief
Lang and Merrikh show that resolution of head-on, but not codirectional, conflicts between replication and transcription machineries requires type II topoisomerases, suggesting that a fundamental difference between the two types of conflicts is supercoil buildup in DNA. Furthermore, they show that supercoil resolution at head-on conflict regions drives R-loop formation.
INTRODUCTION
Transcription and DNA replication occur simultaneously on the same template. The lack of spatiotemporal separation between these two processes leads to conflicts between them multiple times every replication cycle. The replication and transcription machineries can encounter each other either head-on or codirectionally. Codirectional conflicts occur when genes are transcribed on the leading strand whereas head-on conflicts occur when genes are transcribed on the lagging strand. It has been demonstrated that head-on conflicts are more deleterious than codirectional conflicts in that they cause increased mutagenesis, DNA breaks, replisome stalling, and replication restart across diverse organisms (Chappidi et al., 2020; Dutta et al., 2011; French, 1992; Hamperl et al., 2017; Lang et al., 2017; Merrikh and Merrikh, 2018; Merrikh et al., 2011; Million-Weaver et al., 2015a, 2015b; Mirkin and Mirkin, 2005; Paul et al., 2013; Pomerantz and O’Donnell, 2010; Prado and Aguilera, 2005; Wang et al., 2007). Despite many insightful studies into these inevitable encounters, the fundamental question regarding why head-on conflicts are more detrimental than codirectional conflicts remains unanswered. It is perplexing that encounters between the same two machineries (the replication machinery or the replisome and RNA polymerase [RNAP]) can have such different outcomes simply due to orientation.
Topological constraints could explain why head-on conflicts are more deleterious than codirectional conflicts. Unwinding of DNA during transcription generates positively supercoiled DNA ahead and negatively supercoiled DNA behind RNAP (Liu and Wang, 1987; Wu et al., 1988). Similarly, during replication, positive supercoils accumulate in front of the replisome (Hiasa and Marians, 1996; Postow et al., 1999; Vos et al., 2011). The resolution of this supercoiled DNA is critical for both transcription and replication to proceed efficiently (Khodursky et al., 2000). In a codirectional conflict, the positive supercoiling generated in front of the replisome would encounter the negative supercoiling produced from active RNAPs ahead. This would most likely cause a net neutral change in local supercoiling levels. However, during a head-on conflict, the positive supercoiling generated ahead of the replisome would encounter the positive supercoiling produced by RNAP. Therefore, in a head-on conflict, there may be a transient buildup of positive supercoils that has the potential to change the fundamental mechanics of the replisome and RNAP. Such changes could stall the replisome, leading to disassembly and changing the dynamics of RNAP movement and associated mRNAs. These predictions suggest that torsional stress could be the key driver of conflict severity, and therefore, this model must be tested.
Another important question is whether topoisomerases are critical conflict resolution factors. The resolution of supercoils in all organisms requires topoisomerases (Champoux, 2001; Vos et al., 2011; Wang, 2002). In bacteria, there are two topoisomerases that relax positive supercoils, DNA gyrase and Topo IV. DNA gyrase and Topo IV are both required for replication fork progression in vivo (Ashley et al., 2017; Crisona et al., 2000; Khodursky et al., 2000; Peng and Marians, 1993; Vos et al., 2011). Topo IV also plays a critical role in the resolution of catenanes (intertwined chromosomes) as well as the separation of sister chromatids during segregation (Hiasa and Marians, 1996; Zechiedrich and Cozzarelli, 1995). If the torsional stress hypothesis is correct, then type II topoisomerases should be critical conflict resolution factors, yet this question has not been addressed.
Here, we report that type II topoisomerases preferentially associate with head-on genes and that cells harboring engineered head-on conflicts are sensitized to type II topoisomerase inhibitors. Accordingly, we find that conditional depletion of either gyrase or Topo IV is deleterious to cells experiencing engineered head-on conflicts. Inhibition of type II topoisomerase activity leads to increased stalling of the replisome when it approaches a gene transcribed in the head-on, but not codirectional, orientation. Remarkably, however, our data strongly suggest that negative supercoil introduction by DNA gyrase at head-on conflict regions is responsible for the formation of R-loops in these regions. Consistent with this finding, we observe that in cells lacking the RNase HIII enzyme, which resolves R-loops, inhibition of type II topoisomerases lowers R-loop abundance and alleviates R-loop-induced replisome stalling at head-on genes. Furthermore, an allele of gyrase that is strongly defective in the introduction of negative supercoils completely rescues the head-on-conflict-induced lethality of cells lacking RNase HIII. This rescue is also observed when cells are exposed to lysozyme-induced cell wall stress, which is well known to induce a number of endogenous genes, including head-on operons that range from 3 to 6 kb in length (Guariglia-Oropeza and Helmann, 2011).
RESULTS
Type II topoisomerases preferentially associate with a head-on, but not codirectional, engineered conflict region
The relaxation of both positive and negative supercoils is an essential process in all cells. In Bacillus subtilis, relaxation of positive supercoils is accomplished by the activity of either gyrase or Topo IV (Ashley et al., 2017; Crisona et al., 2000; Postow et al., 2001a; Vos et al., 2011). If the model of positive supercoil accumulation at head-on conflict regions is correct, then these enzymes should preferentially associate with a head-on conflict region. To test this hypothesis, we measured gyrase and Topo IV enrichment genome-wide using chromatin immunoprecipitation followed by deep sequencing (ChIP-seq). In order to study the effects of topology at head-on conflict regions, we took advantage of several different tightly controlled engineered conflict systems, all of which were integrated onto the chromosome. In each of these systems, the same exact gene (e.g., lacZ) was inserted onto the same locus in either the head-on or codirectional orientation with respect to replication. To control for gene expression levels, both the head-on and codirectional versions of the gene were placed under the control of the same promoter. In particular, we chose promoters (e.g., Pspank(hy)) that achieve transcrition levels that are close to those of essential and highly transcribed genes that are oriented codirectionally (see Figure S6 for quantification of levels relative to rRNA for the codirectional gene). Such high levels of transcription for the majority of head-on genes are only achieved under specific conditions, such as during exposure to environmental stresses (Guariglia-Oropeza and Helmann, 2011; Lang et al., 2017; Mostertz et al., 2004; Nicolas et al., 2012). Therefore, we did not expect to see enrichment of type II topoisomerases at endogenous head-on genes during growth in rich media. Lastly, our previous data indicated that transcription levels are the same in both orientations in this engineered conflict system (Lang et al., 2017).
In order to measure the relative association of type II topoisomerases with the conflict regions, we used a GFP fusion to the GyrA subunit of gyrase (Tadesse and Graumann, 2006) and constructed a 3xMyc fusion to the ParC subunit of Topo IV. We expressed an IPTG-inducible lacZ gene in either the head-on or codirectional orientation and performed ChIP-seq experiments to obtain a high-resolution map of the association of type II topoisomerases with both the genome and, specifically, the engineered conflict regions.
We found that both gyrase and Topo IV are preferentially enriched at the engineered conflict locus when the orientation of lacZ is head on (Figures 1A, 1B, S1A, S1C, and S1E). Compared to other peaks identified across the genome, the gyrase and Topo IV enrichment at the engineered head-on conflict was one of the largest peaks, suggesting a significant topological problem in the engineered conflict locus (Figures S1B and S1C; Tables S1 and S2). Importantly, this enrichment was transcription dependent. When we measured enrichment of these topoisomerases using ChIP-qPCR, we found that in the absence of the inducer, IPTG, the levels of topoisomerases at the engineered conflict regions were similar in the two orientations (Figures 1C and 1D). Furthermore, we confirmed that the GyrA signal was specific by performing control ChIPs of GFP only (unfused to GyrA) and found no enrichment at the lacZ gene in either orientation (Figure 1E). It is noteworthy that we utilized standard formaldehyde crosslinking for the GyrA ChIPs. However, we were unable to ChIP ParC using formaldehyde. The ParC association was only detectable when we performed the ChIP assays using ciprofloxacin crosslinking, which specifically crosslinks active type II topoisomerases on DNA.
Figure 1. Type II topoisomerases are enriched at head-on genes.
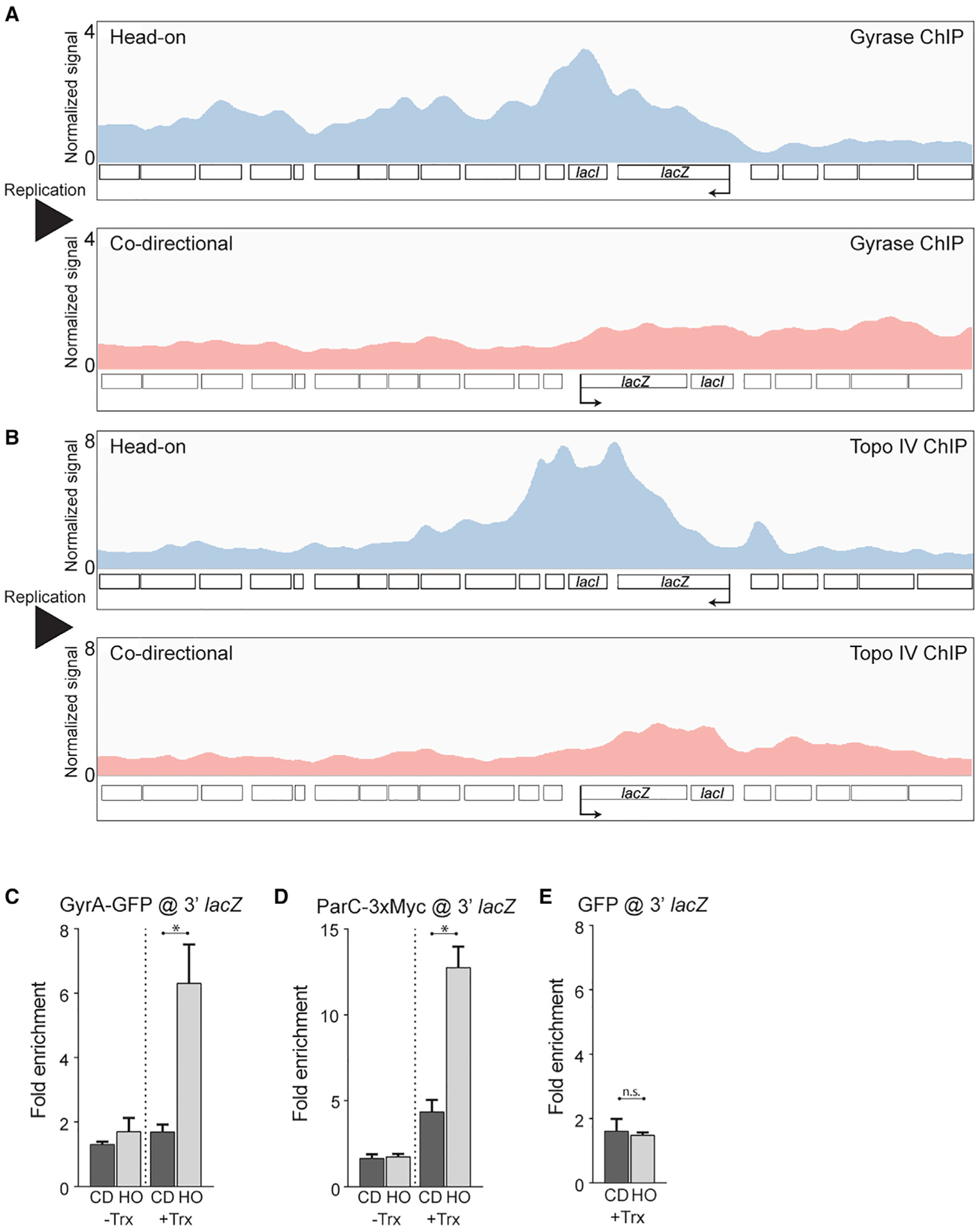
(A and B) DNA gyrase (A) and Topo IV (B) ChIP-seq profiles of cells carrying either a head-on (HO; blue, strain HM3863 [gyrase], HM4074 [ParC]) or codirectional (CD; red, strain HM3864 [gyrase], HM4075 [ParC]) lacZ engineered conflict. Normalized signal is the read depth of immunoprecipitate (IP)/input normalized to the total number of reads. The direction of DNA replication is left to right. Direction of transcription is indicated by the promoter arrow on lacZ.
(C–E) ChIP-qPCR analysis of DNA gyrase (C), Topo IV (D), and GFP (E) in cells carrying either an HO (strain HM3863 [gyrase], HM4074 [ParC], HM3019 [GFP]) or CD (strain HM3864 [gyrase], HM4075 [ParC], HM3020 [GFP]) lacZ engineered conflict. “Trx” refers to transcription of the engineered conflict. Relative enrichment is the signal of lacZ normalized to input relative to a control locus, yhaX, normalized to input. Bars represent the mean and standard error. *p < 0.05; n.s., not significant.
Inhibition of type II topoisomerases increases the association of DnaC (the replicative helicase) at head-on, but not codirectional, genes
In E. coli, gyrase and Topo IV promote replication fork progression (Khodursky et al., 2000). If torsional stress is a major problem at head-on conflict regions, then subtle inhibition of these topoisomerases should lead to increased replication fork stalling at these loci. We tested this hypothesis by performing ChIP-seq of the replicative helicase, DnaC, as a proxy for replication stalling. If fork progression is unimpeded, then the distribution of DnaC enrichment should be equal along the genome in asynchronous bacterial cultures. We have demonstrated previously that DnaC enrichment is a good proxy for replication fork stalling (Lang et al., 2017; Merrikh et al., 2015, 2011). To inhibit type II topoisomerase activity, we used subinhibitory doses of the antibiotic novobiocin. Novobiocin is a competitive inhibitor of type II topoisomerase ATPase activity (Hardy and Cozzarelli, 2003; Maxwell, 1993; Sugino et al., 1978). We performed ChIP-seq experiments in which we measured the association of DnaC genome-wide, which includes the engineered conflict loci in media with and without sublethal concentrations of novobiocin (375 ng/mL). Genome-wide, there are very few DnaC peaks, and those we identified happen to be near the terminus, as previously shown (Tables S2 and S6; Figure S2) (Merrikh et al., 2015; Smits et al., 2011). We found that when the cells were treated with novobiocin, there was an increase in DnaC enrichment at the head-on, but not codirectional, conflict region (Figures 2 and S2; Table S3). These results suggest that without type II topoisomerase activity, topological problems at head-on genes can impede replication.
Figure 2. Type II topoisomerase inhibition results in increased DnaC accumulation at HO genes.
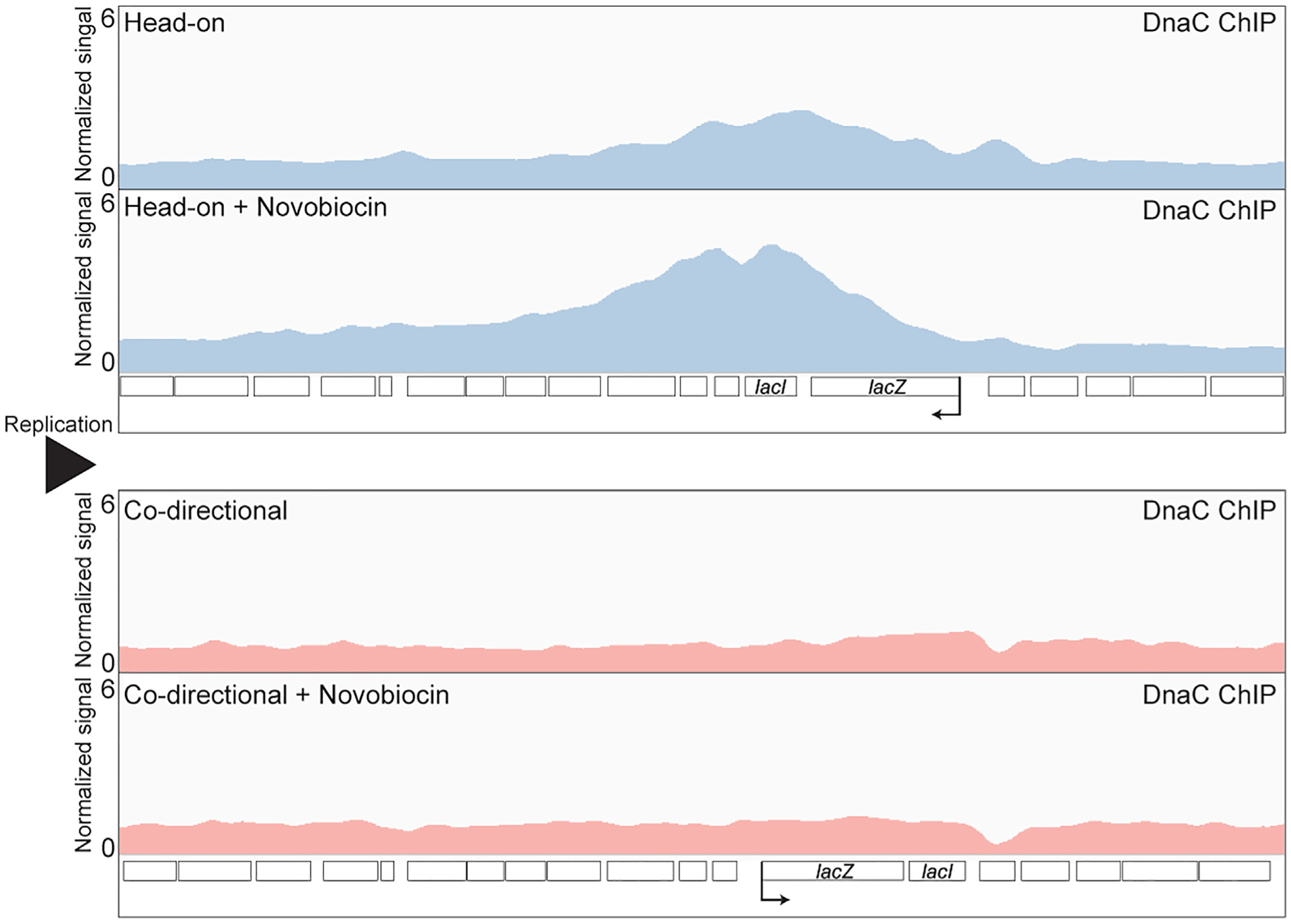
DnaC ChIP-seq profiles of cells carrying either an HO (blue, strain HM3863) or CD (red, strain HM2864) lacZ engineered conflict, with and without novobiocin treatment (375 ng/mL). Normalized signal is the read depth of IP/input normalized to the total number of reads. The direction of DNA replication is left to right. Direction of transcription is indicated by the promoter arrow on lacZ.
Inhibition of type II topoisomerase activity compromises cell survival specifically in the presence of a strong head-on conflict
We previously showed that in the absence of critical conflict resolution factors, head-on conflicts can significantly compromise survival efficiency (Lang et al., 2017; Merrikh et al., 2015; Million-Weaver et al., 2015b). If type II topoisomerases are indeed important for conflict resolution, then the inhibition of these enzymes should impact survival of cells experiencing head-on conflicts. To test this hypothesis, we measured survival efficiency using colony-forming units (CFUs) of cells containing the engineered conflicts in the head-on or codirectional orientation upon chronic treatment with various concentrations of novobiocin. In the absence of novobiocin, there was no difference in survival efficiency of cells containing the engineered conflict in either orientation and regardless of whether the lacZ gene was transcribed (Figure 3A). When the cells were plated on novobiocin, again, there was no difference in survival efficiency between cells carrying the head-on or codirectional lacZ when transcription was off. However, when transcription was turned on, the cells carrying the head-on-, but not codirectionally, oriented lacZ gene were sensitized to low doses of novobiocin. The effects of head-on conflicts on survival, in response to inhibition of type II topoisomerases, was not specific to the chromosomal location or the nature of the gene used to induce the conflict.
Figure 3. DNA gyrase and Topo IV act in parallel to resolve HO conflicts.
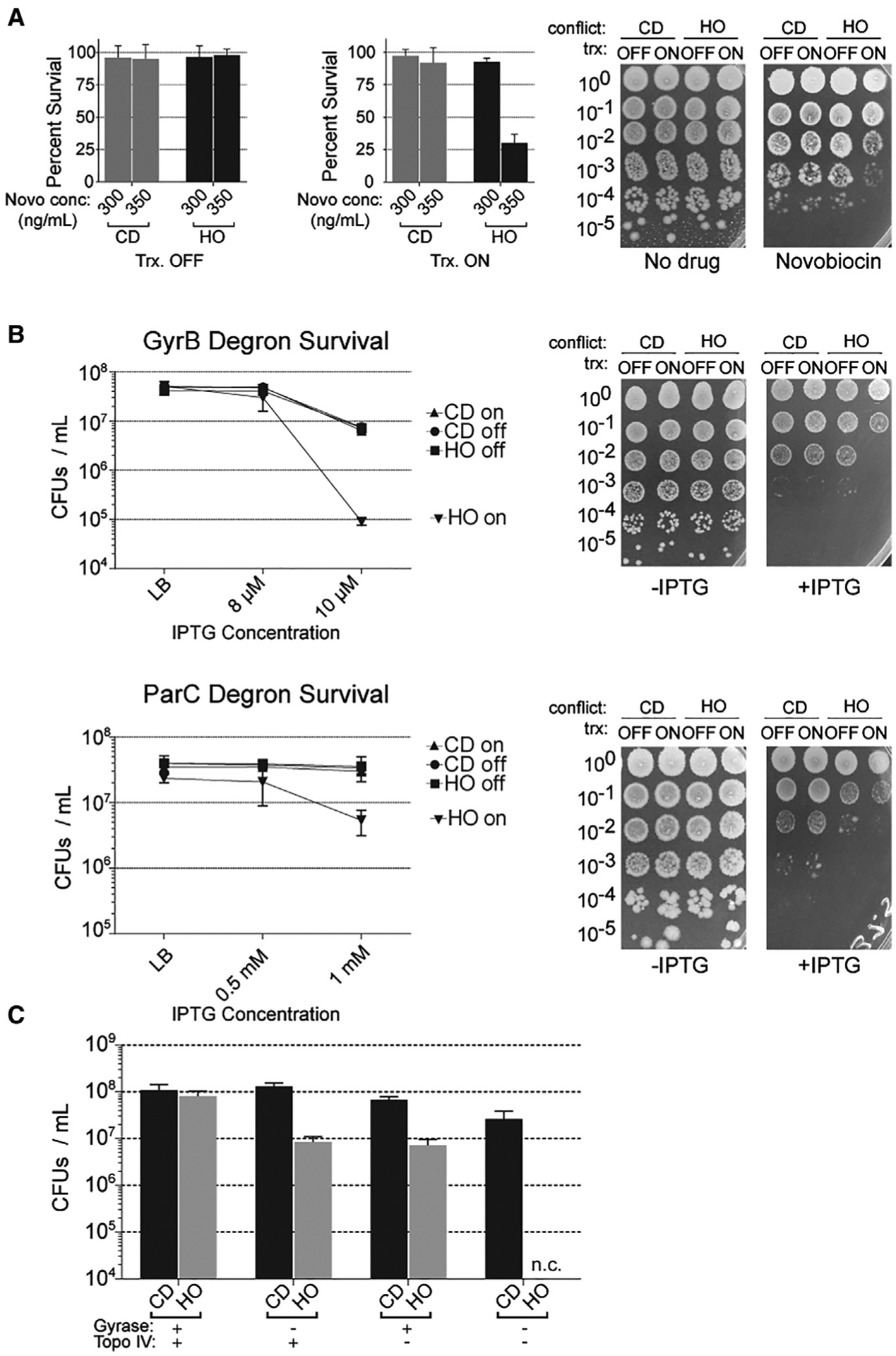
(A) Survival of cells harboring either a repressed (HO, HM640; CD, HM1794) or constitutively transcribed (HO, HM211; CD, HM1795) lacZ engineered conflict plated on LB or LB supplemented with novobiocin (300 or 350 ng/mL). Bar graphs are quantification (mean and standard deviation) of three independent biological replicates.
(B) Survival after conditional (IPTG-dependent) depletion of either gyrase or Topo IV in cells harboring either a repressed (HO, HM1951/HM1467; CD, HM1949/HM1468) or constitutively transcribed (HO, HM1952/HM1450; CD, HM1950/HM1469) lacZ engineered conflict plated on LB or LB supplemented with IPTG (as indicated; plates shown are representative plates of the highest concentration).
(C) Survival of cells harboring a novobiocin-resistant gyrB allele, a conditional gyrase depletion (IPTG-dependent) system and a constitutively transcribed (HO, HM2420; CD, HM2421) lacZ engineered conflict plated on LB or LB supplemented with novobiocin (7 μg/mL), LB supplemented with IPTG (10 μM), or both novobiocin and IPTG.
“Trx” refers to transcription of the engineered conflict. n.c., no countable colonies.
See also Figure S3.
In order to control for potential indirect effects of genomic context, chromosomal location, and sequence, we performed similar survival experiments using a second engineered conflict system. In this system, we inserted a different transcription unit, the luxABCDE operon, onto the opposite (right) arm of the chromosome. We performed the survival experiments with this system as described above. The results of these experiments were consistent with the lacZ system; there was a survival defect in cells containing the luxABCDE operon, but only when this transcription unit was in the head-on orientation and only when the genes were transcribed (Figure S3).
Both gyrase and Topo IV are critical for the resolution of head-on conflicts
Novobiocin has activity against both gyrase and Topo IV, although the affinity of the drug for Topo IV is much weaker than that for gyrase (Peng and Marians, 1993; Sugino et al., 1978). It was unclear from our survival assays whether the survival defects were a result of inhibition of only gyrase or Topo IV or both. It is likely that only gyrase activity is inhibited at the concentrations of novobiocin we used in our experiments. However, it cannot be ruled out that Topo IV activity is also inhibited to some extent under these conditions. To directly determine the contribution of each of the two enzymes to conflict resolution, we adapted a conditional degradation system (Griffith and Grossman, 2008) to specifically deplete the GyrB subunit of gyrase or the ParC subunit of Topo IV. This system is induced by IPTG, and we confirmed the depletion by western blot (Figure S3C). In order to detect potentially subtle differences in survival of our engineered conflict strains, we used concentrations of IPTG that only slightly depleted GyrB and ParC and subtly impacted survival of wild-type cells (gyrase and Topo IV are essential, so a complete depletion cannot be used here). We then tested the survival of cells carrying engineered conflicts under these conditions, but now the engineered conflicts expressed lacZ from a different promoter, Pxis, which is constitutively active. The “transcription off” control for this engineered conflict is achieved through the use of a strain in which this promoter is constitutively off. In both the GyrB and ParC degron systems, we found that without IPTG, there was no difference in survival efficiency (although colonies become smaller, likely due to depleting essential enzymes) in any of the engineered conflict strains. When we specifically depleted GyrB in cells carrying the codirectional conflict, transcription of lacZ made no difference in survival efficiency. In cells carrying the head-on engineered conflict, however, there were significant defects in survival only when the transcription of the engineered conflict was on (Figure 3B). Similarly, when we depleted ParC, there was an ~90% reduction in the number of CFUs when comparing strains with transcriptionally active versus inactive head-on lacZ (Figure 3B).
In order to address whether gyrase and Topo IV act together or in parallel, we constructed a strain that had a mutation in the gyrB gene that conferred a high level of resistance to novobiocin (R138L). In this background, novobiocin treatment can only impact Topo IV. In this same strain, we fused the gyrB gene to the ssrA tag in order to deplete gyrase with our degron system. We found that low concentrations of IPTG (GyrB depletion) or high levels of novobiocin (Topo IV inhibition) both led to a survival defect in the strain carrying the head-on, but not codirectional, conflict (Figure 3C). When we treated cells with both IPTG and novobiocin, the cells expressing the head-on lacZ gene were not viable (Figure 3C). This result indicates that gyrase and Topo IV are the only two factors that can resolve the torsional stress problem at head-on conflict regions.
Inhibition of type II topoisomerases reduces R-loop formation at head-on conflict regions
There is evidence in the literature that topoisomerase activity can influence R-loop formation, at least in vitro and in human cells (Massé and Drolet, 1999; Tuduri et al., 2009). Furthermore, our results described above strongly suggest that DNA topology is a serious problem at head-on conflict regions. Given our prior results that R-loops contribute to many of the detrimental outcomes of head-on conflicts, we decided to investigate whether resolution of head-on conflicts by topoisomerases influence R-loop formation. We tested this hypothesis by directly measuring R-loop levels at the conflict regions in strains lacking RNase HIII (Lang et al., 2017; Ohtani et al., 1999; Randall et al., 2018). We performed DNA-RNA hybrid immunoprecipitations coupled to deep sequencing (DRIP-seq) experiments using the S9.6 antibody, which recognizes RNA:DNA hybrids. We treated our samples in parallel with RNase H in order to ensure specificity for RNA:DNA hybrids and calculated the percent yield in our pull-downs (Figures 4A and S4). Consistent with what we have measured previously using qPCR (Lang et al., 2017), we found more R-loops when the lacZ gene was expressed in the head-on orientation compared to the codirectional orientation (Figures 4A and S4). The DRIP peak at the engineered head-on conflict is the largest peak relative to the others found genome-wide (Figure S4; Table S5). Using a conservative enrichment of 20-fold over the input, we found 16 other prominent peaks around the genome, many of which were near or spanning the most highly transcribed genes (Table S5).
Figure 4. Resolution of HO conflicts by Type II topoisomerases promotes the formation of R-loops.
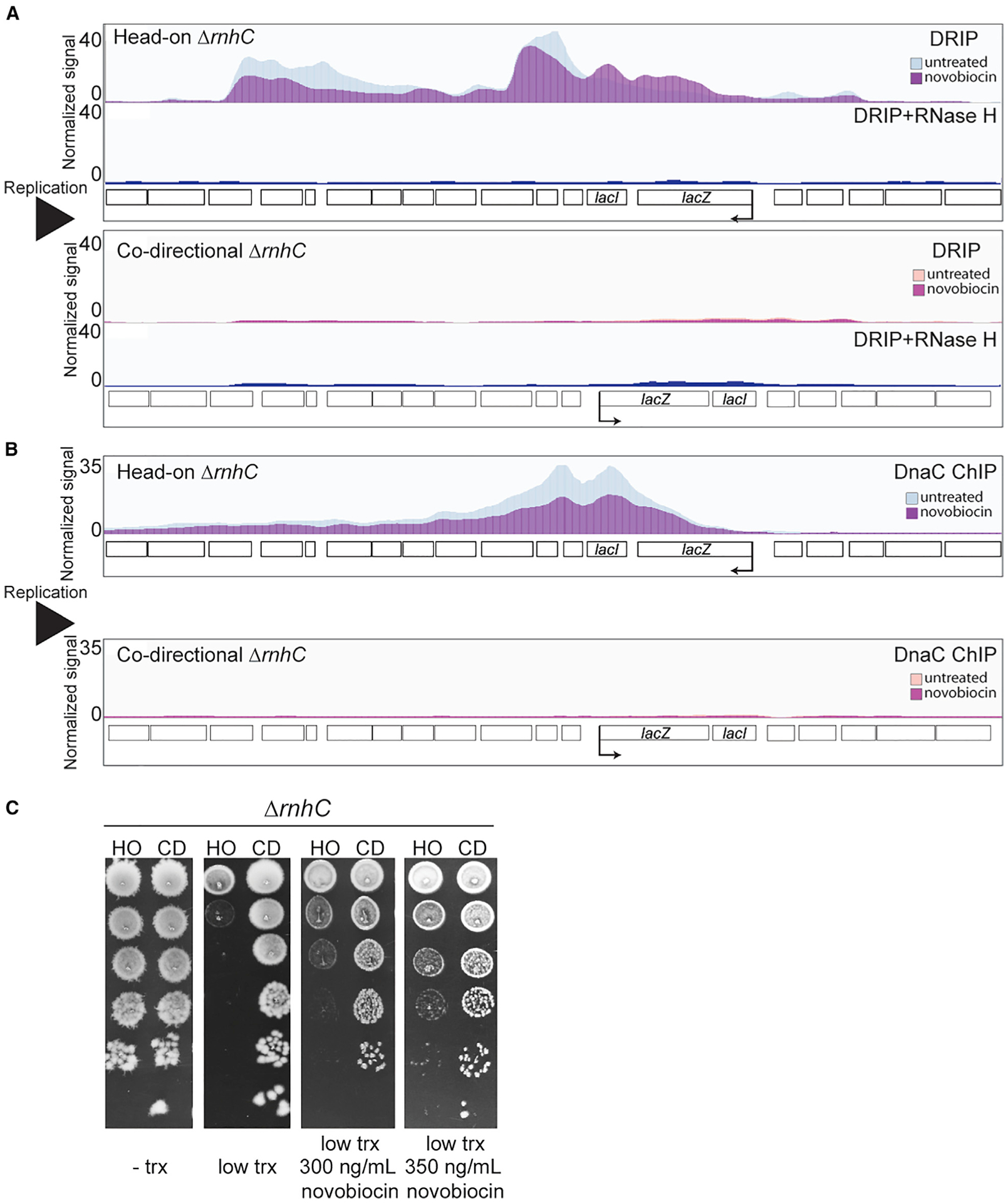
(A and B) DRIP-seq (A) and DnaC (B) ChIP-seq profiles of cells lacking RNase HIII harboring either an HO (blue, strain HM2043) or CD (strain HM2044) lacZ engineered conflict treated or untreated with novobiocin. The bottom panel of both DRIP-seq profiles is the RNase-H-treated control. Normalized signal is the read depth of IP/input normalized to the total number of reads.
(C) Survival of cells lacking RNase HIII harboring either an HO (strain HM2043) or CD (strain HM2044) lacZ engineered conflict treated or untreated with novobiocin.
See also Figures S4 and S5 and Tables S4–S6.
We then used DRIP-seq to measure R-loops in cells treated with low levels of novobiocin to subtly reduce the activities of both type II topoisomerases. Remarkably, we found that when type II topoisomerases are inhibited, R-loop levels are reduced at the head-on conflict region (Figures 4A and S4; Table S5). The lack of a difference in RpoB occupancy at the engineered conflict regions with this amount of novobiocin treatment indicated that the lowered R-loop levels are not simply due to reduced expression of the head-on lacZ gene (Figure S4D). We found that the RpoB signal was much more constrained to the head-on conflict locus compared to the R-loop signal in that region. One interpretation of this unexpected result is that diffusion of supercoils leads to R-loop formation away from the immediate vicinity of the conflict.
Inhibiting type II topoisomerases reduces replisome stalling at the engineered head-on conflict in cells that cannot process R-loops
R-loops at head-on genes stall the replisome in many different organisms (Hamperl et al., 2017; Lang et al., 2017; Prado and Aguilera, 2005). If type II topoisomerase activity is driving R-loop formation at head-on genes, then treating cells with low doses of novobiocin should reduce replisome stalling at head-on conflict regions in cells lacking RNase HIII. We tested this hypothesis using DnaC ChIP-seq, as described above. As we published previously, we found that there is a preferential association of DnaC with head-on versus codirectional conflict regions, and this difference is significantly increased in cells lacking RNase HIII (Figure 4B). This DnaC ChIP signal at head-on conflict regions, in cells lacking RNase HIII, corresponds to complete replication fork stalling at that locus (Lang et al., 2017). When we treated cells with low amounts of novobiocin to inhibit topoisomerase activity, there was a decrease in DnaC enrichment at the head-on conflict region (Figures 4B and S5; Table S6). This result suggests that the type II topoisomerases are responsible for R-loop-mediated replisome stalling at head-on conflict regions.
Inhibiting type II topoisomerases rescues lethality of an engineered head-on conflict in the absence of RNase HIII
We previously showed that increased stalling due to unresolved R-loops at head-on genes is lethal (Lang et al., 2017). If topoisomerase activity is driving R-loop formation at head-on genes, then limiting that activity should increase the viability of cells that contain an engineered head-on conflict and lack RNase HIII. We tested this model by measuring the viability of cells lacking RNase HIII and expressing either the head-on or codirectional lacZ in the presence of low concentrations of novobiocin. As expected, cells with the codirectional engineered conflict had no growth defect when the lacZ gene was induced with IPTG. In contrast, cells expressing the lacZ gene in the head-on orientation had significant cell survival defects. Remarkably, chronic novobiocin exposure rescued these defects in a dose-dependent manner (Figure 4C). Altogether, these results suggest that the resolution of head-on conflicts by type II topoisomerase activity is driving R-loop formation, which affects cell viability.
Introduction of negative supercoils by gyrase promotes R-loop formation at head-on conflict regions
Novobiocin inhibits both gyrase and Topo IV activity. However, gyrase is much more sensitive to novobiocin than Topo IV (Khodursky et al., 2000; Sugino et al., 1978; Levine et al., 1998). We wondered whether the decreased R-loop levels was due to inhibition of gyrase and not inhibition of Topo IV or pleiotropic effects of novobiocin. Gyrase has two activities: (1) relaxation of positive supercoiling and (2) introduction of negative supercoiling (Vos et al., 2011). Both in vitro and in vivo, R-loops have been shown to form more readily (or are more stable) in the presence of gyrase (Drolet et al., 1994, 1995; Massé and Drolet, 1999). This is likely due to the introduction of negative supercoiling by gyrase, as negatively supercoiled DNA will energetically favor R-loop formation, although recent work has suggested that highly positively increased supercoiling could also impact R-loop formation (Stolz et al., 2019). We tested this model by utilizing the gyrB (R138L) mutant, which has reduced ATPase activity and thus has a 10-fold reduction in the ability to introduce negative supercoils (Contreras and Maxwell, 1992; Gross et al., 2003). Whether and/or how much this mutation impacts the positive supercoil relaxation activity of gyrase has not been assessed. However, Topo IV can resolve torsional stress at conflict regions in parallel to gyrase, as we showed above. Therefore, even if the positive supercoil relaxation activity of gyrase is impacted by the R138L mutation, the major effect of this mutation at the conflict region will be a loss of negative supercoil introduction. We used survival assays to measure viability of ΔrnhC strains containing the mutant gyrB in the presence of either the head-on-or codirectionally oriented conflicts. As expected, there was no effect of transcription on the viability of the cells carrying the codirectional engineered conflict. Consistent with our previous work, we found that induction of the engineered conflict was completely lethal when it was oriented head-on to replication. Remarkably, we found that the gyrB R138L mutation completely rescued this lethality (Figure 5A). This rescue was not due to altered transcript levels due to the R138L mutation (Figure S6A). We tested whether this rescue was due to the reduction of R-loops at the conflict region by measuring R-loop association levels directly by DRIP-qPCRs. In cells lacking RNase HIII, consistent with what we have previously reported, we measured roughly 5-fold higher R-loop signal at the head-on compared to the codirectional lacZ (Figure 5B). When we measured R-loops in cells with the R138L gyrB mutation, the R-loop levels were similar at the head-on and codirectional conflict regions. These results demonstrate that it is specifically the introduction of negative supercoils by gyrase at head-on conflict regions that leads to the formation (and/or stability) of R-loops.
Figure 5. DNA gyrase impacts R-loop formation at HO genes.
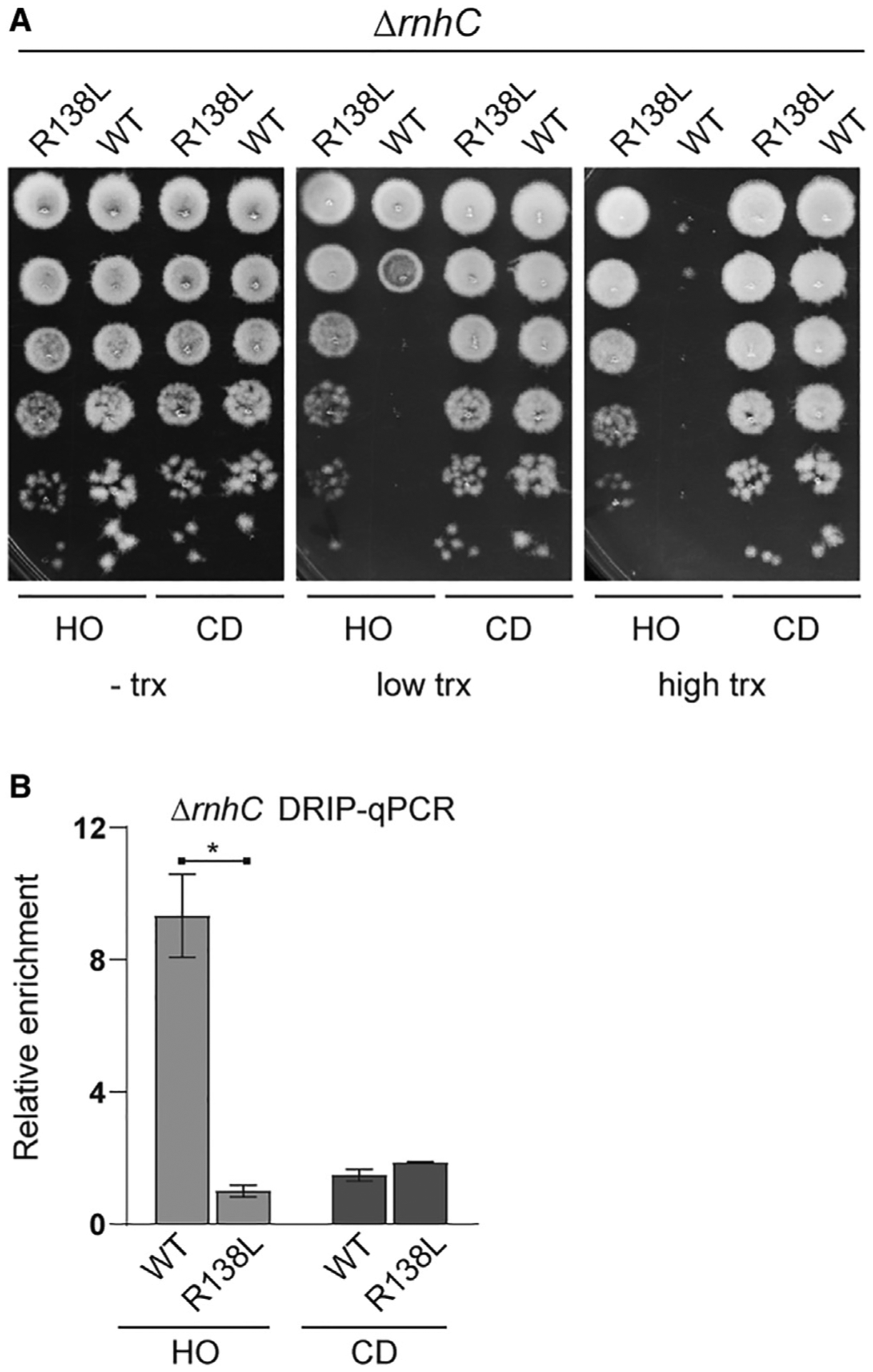
(A) Survival of cells lacking RNase HIII with either the wild-type (WT) or R138L gyrB allele harboring either an HO (HM2043/HM4065) or CD (HM2044/HM4066) lacZ engineered conflict.
(B) DRIP-qPCR analysis of cells lacking RNase HIII with either the WT or R138L gyrB allele harboring either a HO (HM2043/HM4065) or CD (HM2044/HM4066) lacZ engineered conflict. Relative enrichment is the signal of lacZ normalized to input relative to a control locus yhaX normalized to input. Bars represent the mean and standard error of four biological replicates. *p < 0.05.
See also Figure S6.
When we chronically expose cells to various stresses, including cell wall stress induced by lysozyme exposure, we observe a defect in survival of ΔrnhC cells relative to wild-type (Lang et al., 2017). We previously proposed that this phenotype is a result of conflict-induced problems at head-on stress response genes. Here, we tested whether the supercoiling induced R-loop formation is a potential problem at endogenous head-on genes, leading to the observed phenotypic defects. We find that the R138L gyrB mutation suppresses stress response defects of the ΔrnhC strain. We hypothesize that this phenotypic rescue is due to a decrease in R-loop formation at lysozyme resistance genes that are encoded head on (Figure S6B) (see Guariglia-Oropeza and Helmann, 2011 for more information regarding key lysozyme resistance genes). Any gene that responds to lysozyme stress in the codirectional orientation should not experience excess R-loop formation. This is true for all stressors; although stress response genes are encoded in both orientations, only the head-on-oriented ones will experience excess R-loop formation upon induction. Altogether, these results are consistent with the idea that our engineered conflict systems are representative of what occurs at endogenous head-on genes when they are induced. Future experiments should be performed to further investigate the impact of conflicts at endogenous genes, although delineating the impact of various stresses, gene length, and transcription levels will require significant effort.
DISCUSSION
The problem of replication-transcription conflicts exists in all domains of life. Gene-orientation-dependent effects of transcription on DNA replication have been a topic of interest since the analysis of genome organization in E. coli (Brewer, 1988), followed by the discovery that strong head-on transcription slows replication significantly more than codirectional transcription (French, 1992). However, why the orientation of transcription relative to DNA replication matters has remained a mystery. The protein makeup of the two machineries is the same in both orientations, yet the direction in which they encounter each other has profound downstream effects. In this work, we address at least one of the major underlying reasons for this difference.
Our results strongly suggest that positive supercoils build up at head-on conflict regions. We also find that gyrase activity at head-on genes drives R-loop formation. These results can be explained by several potential models. First, a “spin diffusion” model could explain our observations. In this model, excess negative supercoils generated by gyrase promote R-loop formation through the diffusion of the supercoils past RNAPs (Figure 6). This process would be initially triggered by positive supercoil buildup between the replication and transcription machineries at head-on conflict regions, which is rapidly removed by type II topoisomerases. Gyrase would then lead to the generation of hyper-negatively supercoiled DNA (Ashley et al., 2017; Drlica, 1992; Drolet et al., 1994, 1995; Lynch and Wang, 1993). This increase in negative supercoiling would then diffuse through RNAP spinning about its axis (Nudler, 2009, 2012; Ma et al., 2013; Lodge et al., 1989). Alternatively, given that gyrase is recruited over a broad area, it could introduce negative supercoils across the region without a need for RNAP spinning. It has previously been shown that supercoils are constrained when RNAP is unable to spin due to expression of membrane-bound protein-coding gene (Lodge et al., 1989). In a second model, the sudden release of torsional strain by type II topoisomerases could cause RNAP to rapidly progress, generating excessive negative supercoils and R-loop formation (Kuzminov, 2018). Additionally, R-loops could form in front of RNAP due to RNAP backtracking, exposing the 3′ end of the nascent mRNA (Nudler, 2012). This exposed 3′ end could reanneal to the coding DNA strand, forming an R-loop. These models are not mutually exclusive and could all be contributing to R-loop formation and stability at head-on gene regions.
Figure 6. Proposed model for topological changes and R-loop formation at HO conflict regions.
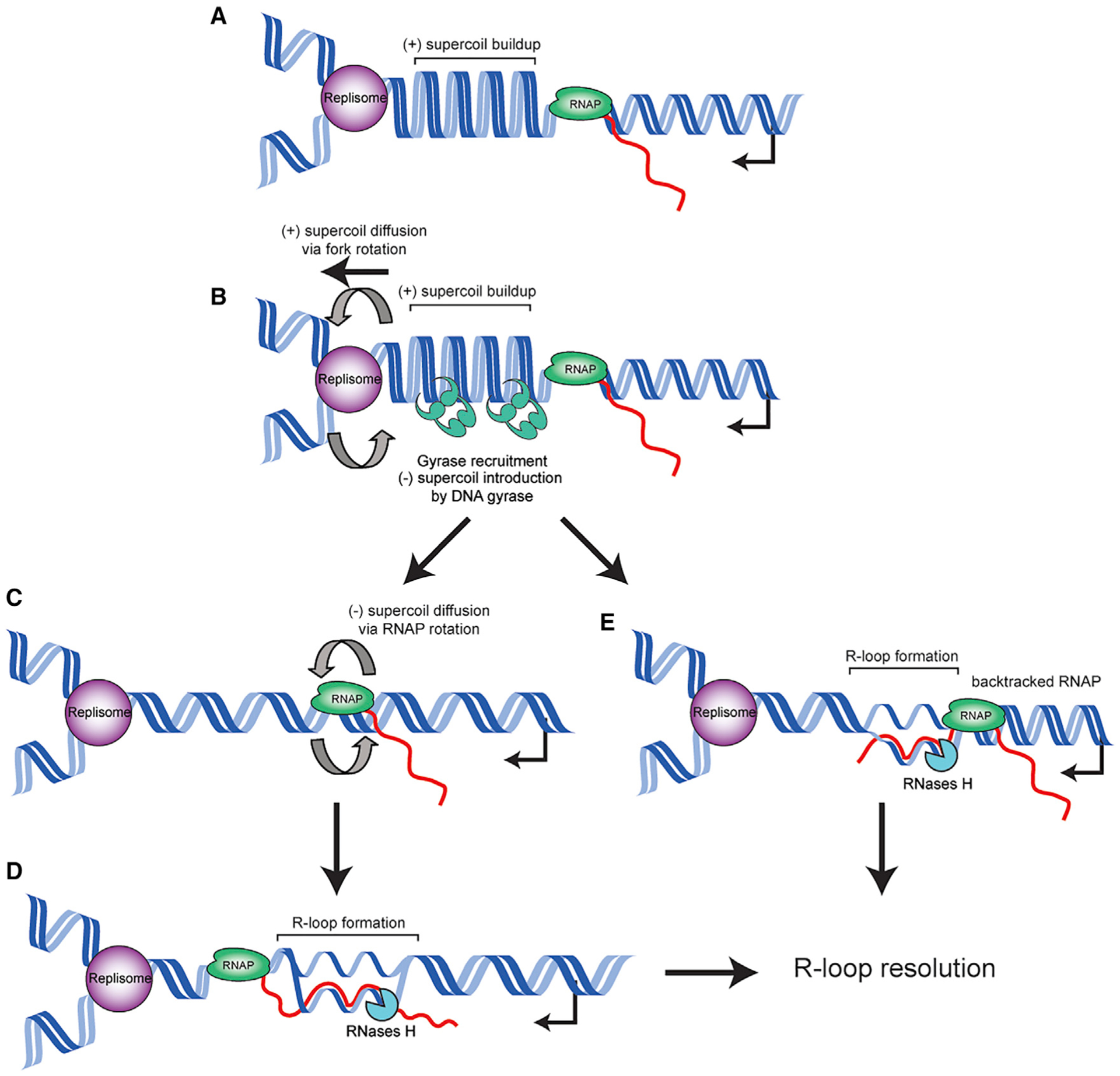
(A) As the replisome and HO transcription unit converge, positive supercoils accumulate in between the two machineries.
(B) DNA gyrase resolves the positive supercoil buildup. The replisome also likely spins to relieve the torsional strain, producing catenanes behind the replication fork, which are resolved by Topo IV.
(C) Gyrase activity rapidly converts the conflict region to negatively supercoiled DNA, causing RNAP to spin about its axis. Negative supercoils diffuse behind RNAP.
(D) The diffused negative supercoils drive R-loop formation behind RNAP, which are resolved by RNase H enzymes.
(E) Alternatively, topological problems cause RNAP to backtrack, allowing an R-loop to form from the exposed 3′ end of the nascent mRNA.
In our previous work, we observed that cells can no longer replicate the chromosome after encountering R-loops at our engineered head-on conflict systems (Lang et al., 2017). One possible explanation for this lethality is that the R-loops completely block the replisome, either because the replicative helicase cannot unwind them or because they stabilize RNAP such that it cannot be removed, resulting in a barrier to the replication fork. Alternatively, many stalled forks in the same genomic region could lead to toxic recombination events and/or production of unresolvable replication intermediates that can be lethal to cells (Magner et al., 2007). Whatever the root cause, the viability of cells with a highly transcribed gene in the head-on orientation requires resolution of R-loops.
The importance of Topo IV in resolving head-on conflicts adds a second dimension to our findings. The observations that Topo IV is important for conflict resolution can be interpreted in two ways: (1) Topo IV helps relax positive supercoils at conflict regions, and/or (2) the increased torsional stress leads to the formation of catenanes by inducing replisome spinning about its axis (Bermejo et al., 2007; Keszthelyi et al., 2016; Schalbetter et al., 2015). Given that there is a significant amount of literature showing that Topo IV is critical for catenane resolution, we favor the second possibility (Espeli and Marians, 2004; Levine et al., 1998; Zechiedrich and Cozzarelli, 1995). These models, however, are not mutually exclusive.
In addition to the topology model, one hypothesis that could explain gene orientation effects of conflicts is the strand specificity of where the replicative helicase resides (lagging strand in bacteria and leading strand in eukaryotes) (Gómez-González and Aguilera, 2019; Hamperl and Cimprich, 2016). This model could explain why the two different types of conflicts have differential consequences. However, the discovery that R-loops are a major problem in head-on, but not codirectional, conflicts in both bacteria and mammalian cells undermines this model (Hamperl et al., 2017; Lang et al., 2017). The replicative helicase moves on the lagging strand in bacteria whereas it moves on the leading strand in mammalian cells, yet the fundamental problem of R-loop enrichment in head-on conflicts remains the same across these species. Therefore, gene-orientation-specific problems are unlikely to stem from this particular architectural feature of the replisome complex. On the other hand, production of positive supercoils by the replication and transcription machineries is a universal feature and therefore could be the fundamental mechanism underlying gene-orientation-specific effects of replication-transcription conflicts. Recent work in human cells suggested that topology plays a role in R-loop formation at head-on gene regions, demonstrating the conservation of the results presented here (Promonet et al., 2020).
It is clear from this work, as well as that of others, that after some encounters with the transcription machinery, replication stalls, the replisome collapses, and replication progression requires restart proteins (Mangiameli et al., 2017; Merrikh et al., 2011). However, the extent to which the fork is remodeled and whether there is replication fork reversal after a head-on conflict are not yet clear. Previous studies have implied that in head-on conflicts, the replication fork reverses and is subsequently processed by recombination proteins (Chappidi et al., 2020; De Septenville et al., 2012; Million-Weaver et al., 2015b). Furthermore, it has been shown in vitro that replication forks reverse in response to positive supercoil accumulation (Postow et al., 2001b). Given that at least in eukaryotic systems supercoiling can push the fork back, our data are consistent with the model that conflicts lead to replication fork reversal due to positive supercoil buildup.
We previously proposed that the head-on orientation is retained for some genes as a mechanism to increase mutagenesis and promote gene-specific evolution (Merrikh and Merrikh, 2018; Paul et al., 2013). Further work showed that the increased mutagenesis of head-on genes is driven by R-loops in wild-type cells (Lang et al., 2017). Given that gyrase activity is facilitating R-loop formation, our results suggest that the activity of this enzyme, albeit indirectly, leads to increased mutagenesis. Interestingly, as our group and others have shown, the full capacity of gyrase to introduce negative supercoils is not essential for viability (Gross et al., 2003). Why then is this function conserved? We speculate that the introduction of negative supercoils by gyrase is evolutionarily beneficial. In particular, we previously showed that head-on genes, including many of the critical stress response genes (which are functionally enriched in the head-on orientation), evolve faster than codirectional genes. Under selection, these head-on genes will be highly transcribed, gaining beneficial mutations faster than if they were codirectionally oriented, simply due to a conflict-induced increase in mutation rates. If those beneficial mutations are obtained through negative supercoil introduction by gyrase (and downstream R-loop formation), then this property of gyrase would be retained over evolutionary time despite the fact that it is not immediately necessary for viability. In other words, the activity of gyrase to introduce negative supercoils would hitchhike along in cells that have rapidly adapted to their environment by obtaining beneficial mutations relatively quickly through this mechanism.
In this work, we discovered (what appears to be) the main source of gene-orientation-specific problems in replication-transcription conflicts. We also unraveled an intriguing feature of topoisomerases that, in the big picture, could place them into a category of evolutionarily beneficial factors that increase mutagenesis. These findings highlight the fundamental importance and influence of conflicts and DNA supercoiling on cellular physiology, genome organization, and adaptation.
STAR★METHODS
RESOURCE AVAILABILITY
Lead contact
Further information and requests for resources and reagents should be directed to the Lead Contact, Houra Merrikh (houra.merrikh@vanderbilt.edu).
Materials availability
Materials generated in this study are available upon request.
Data and code availability
Datasets generated during this study are available from NCBI SRA project ID PRJNA691533.
EXPERIMENTAL MODEL AND SUBJECT DETAILS
Bacterial strains and growth conditions
Strains are listed in the key resources table. All strains were constructed in the HM1 (JH642) (Brehm et al., 1973) B. subtilis background. The rnhC::mls mutant (HM711) was obtained from the Bacillus genetic stock center (Columbus, OH). To move the rnhC::mls allele, genomic DNA was extracted from HM711 using a commercially available kit (Thermo) and used to transform into HM1 (and its derivatives with engineered conflict constructs) as per standard protocol (Harwood et al., 1990). Strains were streaked on LB agar plates and supplemented with antibiotics where appropriate. Precultures were inoculated from single colonies into 2 or 5 mL of LB broth and incubated at 37°C with shaking (260 RPM). Precultures were used to inoculate experimental cultures which were grown and treated as indicated for each different experiment in the materials and methods.
KEY RESOURCES TABLE
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| Mouse monoclonal S9.6 DNA:RNA Hybrid antibody | Millipore | MABE1095 |
| Rabbit polyclonal anti-DnaC Antibody | Smits et al., 2010 | N/A |
| Rabbit polyclonal anti-Gfp antibody | Merrikh et al., 2015 | N/A |
| Mouse monoclonal anti-Myc antibody (clone 9E10) | Invitrogen | 13–2500 |
| Mouse monoclonal anti-RpoB antibody (clone 8RB13) | Thermo | MA125425 |
| Bacterial and virus strains | ||
| B. subtilis phe trp | Brehm et al., 1973 | HM1 |
| B. subtilis phe trp thrC::Pxis-lacZ (HO) ICEBs1(0) | Merrikh et al., 2015 | HM211 |
| B. subtilis phe trp thrC::Pxis-lacZ (HO) | Merrikh et al., 2015 | HM640 |
| B. subtilis phe trp amyE::Pspank(hy)-lacZ (HO) | Lang et al., 2017 | HM1300 |
| B. subtilis phe trp amyE::Pspank(hy)-lacZ (CD) | Lang et al., 2017 | HM1416 |
| B. subtilis phe trp thrC::Pxis-lacZ (HO) ICEBs1(0) amyE::Pspank(hy)-sspB parC::parC-ssrA | This study | HM1450 |
| B. subtilis phe trp thrC::Pxis-lacZ (HO) amyE::Pspank(hy)-sspB parC::parC-ssrA | This study | HM1467 |
| B. subtilis phe trp thrC::Pxis-lacZ (CD) amyE::Pspank(hy)-sspB parC::parC-ssrA | This study | HM1468 |
| B. subtilis phe trp thrC::Pxis-lacZ (CD) ICEBs1(0) amyE::Pspank(hy)-sspB parC::parC-ssrA | This study | HM1469 |
| B. subtilis phe trp thrC::Pxis-lacZ (CD) amyE::Pspank(hy)-sspB gyrB::gyrB-ssrA | This study | HM1949 |
| B. subtilis phe trp thrC::Pxis-lacZ (CD) ICEBs1(0) amyE::Pspank(hy)-sspB gyrB::gyrB-ssrA | This study | HM1950 |
| B. subtilis phe trp thrC::Pxis-lacZ (HO) amyE::Pspank(hy)-sspB gyrB::gyrB-ssrA | This study | HM1951 |
| B. subtilis phe trp thrC::Pxis-lacZ (HO) ICEBs1(0) amyE::Pspank(hy)-sspB gyrB::gyrB-ssrA | This study | HM1952 |
| B. subtilis phe trp amyE::Pspank(hy)-lacZ (HO) ΔrnhC::MLS | Lang et al., 2017 | HM2043 |
| B. subtilis phe trp amyE::Pspank(hy)-lacZ (CD) ΔrnhC::MLS | Lang et al., 2017 | HM2044 |
| B. subtilis phe trp thrC::Pxis-lacZ (HO) ICEBs1(0) amyE::Pspank(hy)-sspB gyrB::gyrB(R138L)-ssrA | This study | HM2420 |
| B. subtilis phe trp thrC::Pxis-lacZ (CD) ICEBs1(0) amyE::Pspank(hy)-sspBgyrB::gyrB(R138L)-ssrA | This study | HM2421 |
| B. subtilis phe trp thrC::Pxis-lacZ (HO) ICEBs1(0) amyE::Pspank(hy)-sspBgyrB::gyrB-myc-ssrA | This study | HM2442 |
| B. subtilis phe trp thrC::Pxis-lacZ (HO) ICEBs1(0) amyE::Pspank(hy)-sspB gyrB::parC-myc-ssrA | This study | HM2444 |
| B. subtilis phe trp ΔrnhC::MLS | Lang et al., 2017 | HM2655 |
| B. subtilis amyE::Pspank(hy)-lacZ (HO) lacA::Pspank(hy)-3xmyc-gfp | This study | HM3019 |
| B. subtilis amyE::Pspank(hy)-lacZ (CD) lacA::Pspank(hy)-3xmyc-gfp | This study | HM3020 |
| B. subtilis phe trp amyE::Pspank(hy)-lacZ (HO) gyrA::gyrA-gfp | This study | HM3863 |
| B. subtilis phe trp amyE::Pspank(hy)-lacZ (CD) gyrA::gyrA-gfp | This study | HM3864 |
| B. subtilis phe trp gyrB(R138L) | Samadpour and Merrikh, 2018 | HM3387 |
| B. subtilis phe trp ΔrnhC::MLS gyrB(R138L) | This study | HM4064 |
| B. subtilis phe trp ΔrnhC::MLS gyrB(R138L) amyE::Pspank(hy)-lacZ (HO) | This study | HM4065 |
| B. subtilis phe trp ΔrnhC::MLS gyrB(R138L) amyE::Pspank(hy)-lacZ (CD) | This study | HM4066 |
| B. subtilis phe trp amyE::Pspank(hy)-lacZ (HO) parC::parC-3xMyc | This study | HM4074 |
| B. subtilis phe trp amyE::Pspank(hy)-lacZ (CD) parC::parC-3xMyc | This study | HM4075 |
| Chemicals, peptides, and recombinant proteins | ||
| EcoRV-HF | NEB | R0195 |
| HindIII-HF | NEB | R0104 |
| EcoRI-HF | NEB | R0101 |
| DraI | NEB | R0129 |
| RNase H | NEB | M0297 |
| Protein A Sepharose | GE | GE17-0780-01 |
| Critical commercial assays | ||
| Nextera XT DNA Library Preparation Kit | Illumina | FC-131–1024 |
| NEBNext DNA Library Prep master Mix Set | NEB | E6040 |
| GeneJET PCR Purification Kit | Thermo | K0701 |
| iScript supermix | Bio-Rad | 1708840 |
| iTaq Universal SYBR Green master mix | Bio-Rad | 1725121 |
| GeneJet Genomic DNA purication Kit | Thermo | K0721 |
| Deposited data | ||
| All sequencing DATA uploaded to the Sequence Read Archive | NCBI SRA | Bioproject PRJNA691533 |
| Oligonucleotides | ||
| GACATCCTCTGACAATCCTAGAG | This study | HM86 |
| GGCAGTCACCTTAGAGTGCCCAAC | This study | HM87 |
| GGCTTTCGCTACCTGGAGAG | Lang et al., 2017 | HM188 |
| GACGAAGCCGCCCTGTAAAC | Lang et al., 2017 | HM189 |
| CCGTCTGACCCGATCTTTTA | Lang et al., 2017 | HM192 |
| GTCATGCTGAATGTCGTGCT | Lang et al., 2017 | HM193 |
| AAGGCACATGGCTGAATATCG | Lang et al., 2017 | HM910 |
| ACACCAGACCAACTGGTAATGG | Lang et al., 2017 | HM911 |
| TTATGGATCCTGAAGGGTGAAGATGAACTG | This study | HM1690 |
| TTATTCTAGATTGTTCTGTATGAAGGCGCCAAAC | This study | HM1691 |
| ttatgaattcTATCGTAGAGGGTGACTCTG | This study | HM2282 |
| TTATTCTAGAGATGTCAAGATTTTTAACGTATCTC | This study | HM2283 |
| Recombinant DNA | ||
| pGCS::parC | This study | pHM186 |
| pGCS::gyrB | This study | pHM260 |
| Software and algorithms | ||
| SAMtools | Li et al., 2009 | http://www.htslib.org/ |
| Bowtie 2 | Langmead and Salzberg, 2012 | http://bowtie-bio.sourceforge.net/bowtie2/index.shtml |
| Prism 7 | Graphpad | https://www.graphpad.com/scientific-software/prism/ |
| DeepTools | Ramírez et al., 2014 | https://deeptools.readthedocs.io/en/develop/ |
| IGV | Robinson et al., 2011 | http://software.broadinstitute.org/software/igv/ |
| HOMER | Heinz et al., 2010 | http://homer.ucsd.edu/homer/ |
E. coli DH5α was used to propagate recombinant DNA vectors. Transformations were done using heat shock of competent E. coli. E. coli cultures were grown at 37°C with shaking (260 RPM) in LB supplemented with 50 μg/mL carbenicillin where appropriate. All plasmid vectors were purified using a commercially available plasmid extraction kit (Thermo).
Plasmid and strain constructions
pHM186 PCR was used to amplify 500 bp of the 3′ end of parC without the stop codon (primers HM1690/1691). The resulting amplicon was digested with BamHI and XbaI and ligated into pGCS (Griffith and Grossman, 2008).
pHM260 PCR was used to amplify 500 bp of the 3′ end of gyrB without the stop codon (primers HM22832284). The resulting amplicon was digested with EcoRI and XbaI and ligated into pGCS.
HM1450 Strain HM867 (Merrikh et al., 2015) was transformed with plasmid pHM186 and transformants were selected on LB plates containing chloramphenicol.
HM1467 Strain HM866 (Merrikh et al., 2015) was transformed with plasmid pHM186 and transformants were selected on LB plates containing chloramphenicol.
HM1468 Strain HM868 (Merrikh et al., 2015) was transformed with plasmid pHM186 and transformants were selected on LB plates containing chloramphenicol.
HM1469 Strain HM869 (Merrikh et al., 2015) was transformed with plasmid pHM186 and transformants were selected on LB plates containing chloramphenicol.
HM1949 Strain HM868 was transformed with plasmid pHM190 and transformants were selected on LB plates containing chloramphenicol.
HM1950 Strain HM869 was transformed with plasmid pHM190 and transformants were selected on LB plates containing chloramphenicol.
HM1951 Strain HM866 was transformed with plasmid pHM190 and transformants were selected on LB plates containing chloramphenicol.
HM1952 Strain HM867 was transformed with plasmid pHM190 and transformants were selected on LB plates containing chloramphenicol.
HM2420 Strain HM866 was transformed with genomic DNA purified from HM3387 and transformants were selected on LB plates containing novobiocin (4 μg/mL). The novobiocin resistant transformant was then transformed with pHM260.
HM2421 Strain HM869 was transformed with genomic DNA purified from HM3387 and transformants were selected on LB plates containing novobiocin (4 μg/mL). The novobiocin resistant transformant was then transformed with pHM260.
HM4064 Strain HM3387 was transformed with gDNA purified from strain HM2655 and transformants were selected for on LB containing erythromycin and lincomycin.
HM4065 Strain HM4064 was transformed with plasmid pHM171 (Lang et al., 2017).
HM4066 Strain HM4064 was transformed with plasmid pHM180 (Lang et al., 2017).
METHOD DETAILS
Viability assays
Strains were struck on LB plates supplemented with the appropriate antibiotic from freezer stocks and incubated overnight at 37°C Single colonies were used to inoculate 2 mL LB cultures in glass tubes. The cultures were grown at 37°C with shaking (260 RPM) to OD600 = 0.5–1.0. Precultures were adjusted to OD 0.3 and then serially diluted in 1x Spizzen’s Salts (15 mM ammonium sulfate, 80 mM dibasic potassium phosphate, 44 mM monobasic potassium phosphate, 3.4 mM trisodium citrate, and 0.8 mM magnesium sulfate). 5ul of each dilution was plated onto LB plates and incubated at 30°C overnight. For survival assays with the engineered conflict strains, LB plates were either supplemented or not with various concentrations of novobiocin and/or IPTG as indicated in the figure legends. For the type II topoisomerase degron experiments, chloramphenicol was added to the media to maintain the stability of degron tag. For chronic cell wall stress assays, LB plates were supplemented with lysozyme to a final concentration of 50 μg/mL. Plates were imaged with a BioRad Gel Doc™ XR+ Molecular Imagerâ and colonies were enumerated.
Slot blot analysis
Precultures grown from single colonies were diluted back to OD = 0.05 in replicate cultures and grown until OD600 = 0.3. IPTG was added to one replicate of each strain to a final concentration of 0.1 mM. For each strain, 3 mL of culture was spun at 10k RPM for 3 min and washed with 1x PBS. Cell pellets were resuspended in 300 μL Lysis Buffer (TE pH 8.0, 0.1 mg/mL lysozyme, and 1x AEBSF) and incubated at 37°C for 30 min. Cells were lysed by the addition of SDS to a final concentration of 1%. Samples were then boiled for 10 minutes. Total protein levels were measured using a Qubit protein quantification assay. 40 μg of each sample was applied to a PVDF membrane via a Slot Blot apparatus (Bio-Rad). Membrane were then blocked in Odyssey Buffer, and anti-myc antibody (910E1, invitrogen) was added (1:500) for overnight incubation at 4°C. Membranes were washed 5x in PBST. Membranes were then incubated with an anti-mouse Odyssey secondary antibody (1:15,000 in Odyssey Buffer) for 30 mins. Membranes were then washed 3x in PBST and imaged on a Li-Cor Imager.
Chromatin immunoprecipitation assays (ChIPs)
Precultures were diluted to OD600 of 0.05 in LB and grown at 30°C with shaking. At OD600 ~0.1, cultures were induced with 1 mM IPTG (final concentration) and grown until the culture was at OD600 = 0.3 and processed as described (Merrikh et al., 2011). Briefly, cultures were crosslinked with 1% formaldehyde or ciprofloxacin (4 ug/mL, Topo IV only) for 20 minutes and subsequently quenched with 0.5 M glycine (formaldehyde crosslinking only). Cell pellets were collected by centrifugation and washed once with cold phosphate buffered saline (PBS). Cell pellets were resuspended with 1.5 mL of Solution A (10 mM Tris–HCl pH 8.0, 20% w/v sucrose, 50 mM NaCl, 10 mM EDTA, 10 mg/ml lysozyme, 1 mM AEBSF) and incubated at 37°C for 30 min. After incubation, 1.5 mL of 2x IP buffer (100 mM Tris pH 7.0, 10 mM EDTA, 20% triton x-100, 300 mM NaCl and 1mM AEBSF) was added and lysates were incubated on ice for 30 minutes. Lysates were then sonicated 4 times at 30% amplitude for 10 s of sonication and 10 s of rest. Lysates were pelleted by centrifugation at 8000 RPMs for 15 minutes at 4°C. Each IP was done with 1 mL of cell lysate and 40 μL was taken out prior to addition of the antibody as an input control. IPs were performed using rabbit polyclonal antibodies against DnaC (Smits et al., 2010), RNAP (Santa Cruz Biotech), GFP (Abcam, gyrase) and Myc (Invitrogen, Topo IV). IPs were rotated overnight at 4°C. After incubation with the antibody, 30 μL of 50% Protein A Sepharose beads (GE) were added and IPs were incubated at RT for one hour with gentle rotation. Beads were then pelleted by centrifugation at 2000 RPM for 1 minute. The supernatant was removed and the beads were washed 6x with 1mL of 1x IP buffer. An additional wash was done with 1 mL of TE pH 8.0. After the washes, 100 μL of elution buffer I (50 mM Tris pH 8.0, 10 mM EDTA, 1% SDS) was added and beads were incubated at 65°C for 10 minutes. Beads were pelleted by centrifugation at 5000 RPMs for 1 minute. The supernatant was removed, saved and 150 μL of elution buffer II (10 mM Tris pH 8.0, 1 mM EDTA, 0.67% SDS) was added. Beads were then pelleted by centrifugation at 7000 RPMs for 1 minute and the supernatant was combined with the first elution. The combined eluates were then de-crosslinked by incubation at 65°C for overnight. The eluates were then treated with proteinase K (0.4 mg/mL) at 37°C for 2 hours. DNA was then extracted with a GeneJet PCR purification Kit (Thermo) according to the manufacturer’s instructions or a standard phenol:chloroform extraction.
DNA:RNA hybrid immunoprecipitation assays (DRIPs)
DRIPs were performed as described with modifications for use in bacteria(García-Rubio et al., 2018; Lang et al., 2017; Sanz and Chédin, 2019). Precultures were diluted to OD600 of 0.05 in LB and grown at 30°C with shaking. At OD600 ~0.1, cultures were induced with 1 mM IPTG (final concentration) and grown until the culture was at OD600 = 0.3. Cells were pelleted by centrifugation and washed twice with cold PBS. Total nucleic acids were purified from cell pellets using phenol:chloroform extraction and ethanol precipitation. Precipitated DNA was spooled on a glass rod and after drying, DNA was resuspended in TE pH 8.0 and treated with EcoRV, EcoRI, DraI, and HindIII overnight at 37°C. Cutsites in the engineered locus (and ~20 kb window surrounding) are listed in Table S4. Digested chromosomal DNA was then purified by phenol:chloroform extraction and brought to final volume of 125 μL. For the RNase H treated controls, 10 μg was treated with 3 μL of RNase H (NEB) in 1x RNase H buffer at 37°C overnight prior to immunoprecipitation. Nucleic acids were then quantified using a Qubit (Invitrogen) and 10 μg were added to each IP in 500 total μL of TE. 50 μL was then removed kept as INPUT. 52 μL of 10x Binding buffer (100 mM NaPO4 pH 7.0, 1.4 M NaCl, 0.5% Triton X-100) was added. S9.6 antibody (Millipore) was added (20 μL) and samples were incubated overnight at 4°C with gentle rotation. After incubation with the antibody, 40 μL of 50% Protein A Sepharose beads (GE) were added and IPs were incubated at 4°C for 2 hours with gentle rotation. Beads were then pelleted by centrifugation at 2000 RPM for 1 minute. The supernatant was removed and the beads were washed 3x with 1mL of 1x Binding buffer. After the washes, 300 μL of elution buffer (10 mM Tris pH 8.0, 1 mM EDTA, 0.67% SDS) and 7 μL Proteinase K (QIAGEN) were added. For the INPUT samples, 3 μL Proteinase K was added. All samples were incubated at 55°C for 45 minutes with gentle rotation. Beads were then pelleted by centrifugation at 7000 RPMs for 1 minute and the supernatant moved to a new tube. DNA was purified by phenol:chloroform extraction and ethanol precipitation and used to prepare Illumina libraries using the Nextera NT library prep kit (Illumina) or the NEBNext Library Prep Kit (NEB) or analyzed using qPCR. DRIP-qPCR analysis was done by the ratio of signal at the conflict region (primer pair 188/189 or 910/911) divided by a control locus yhaX (192/193).
RNA extraction and cDNA preparation
Cells were grown in LB to mid-exponential phase and back diluted to OD600 0.05 into LB either supplemented with or lacking 1mM IPTG. Cells were grown for 2 hours at 30°C (3 generations) prior to harvesting. 5 mL of culture was harvested by addition to an equal volume of ice-cold methanol followed by centrifugation at 4,000×g for 5 minutes. Cells were lysed with 20 μg/mL lysozyme for 10 minutes. RNA was isolated with the GeneJET RNA Purification Kit (ThermoFisher Scientific). 1 μg of RNA was treated with RNase-free DNase I (ThermoFisher Scientific) for 40 minutes at 37°C. DNase I was denatured by the addition of 1ul of EDTA and incubation at 65°C for 10 minutes. Reverse transcription was performed with iScript Supermix (BioRad) as per manufacturer’s instructions. mRNA abundance was measured via qPCR analysis by measuring the signal ratio of the target locus lacZ (primer pair 188/189) by the control rrn locus (primer pair 86/87).
Next generation sequence analysis
Sequencing libraries were generated using either the Nextera NT library prep kit from Illumina or by standard end polishing and ligation with the NEBNext adaptor kit (NEB). Approximately 4M × 150 bp paired-end or single-end Illumina Next-Seq reads per sample were quality and adaptor trimmed using Trimmomatic (Bolger et al., 2014) and mapped to the genome of B. subtilis strains HM1300 (head-on lacZ) and HM1416 (co-directional lacZ) in the strain background JH642 (GenBank: CP007800.1) using Bowtie 2 (Langmead and Salzberg, 2012). Bam files were normalized for the total number of mapped reads and the ratio of the immunoprecipitation versus the input was done using the deepTools bamCompare tool (Ramírez et al., 2014). Plots were generated in IGV (Robinson et al., 2011). Peaks were identified and analyzed using the HOMER (Heinz et al., 2010) findPeaks tool, analyzing the mapped read files for the IP compared to the input. Read density in the identified peak regions was quantified using HOMER’s annotatePeaks function using the normalized bedgraph files (IP/input) generated from deepTools.
QUANTIFICATION AND STATISTICAL ANALYSIS
Analysis of deep sequencing data was done using HOMER as described in Method details. Statistical analysis of ChIP-qPCR and RT-qPCR data was done in Prism 8 as described in Method details.
Supplementary Material
Highlights.
Type II topoisomerases act at head-on replication-transcription conflict regions
Cells with severe head-on conflicts are sensitive to the inhibition of topoisomerases
Type II topoisomerase inhibition increases replisome stalling at head-on genes
Supercoil resolution at head-on conflict regions drives R-loop formation
ACKNOWLEDGMENTS
We would like to thank Christopher Merrikh and the other members (past and present) of the Merrikh lab, as well as Neil Osheroff and Felipe Cortés Ledesma for helpful discussions. The authors would also like to thank Sara Mangiameli for revising the manuscript and Peter Graumann for sharing strains. This work was supported by National Institutes of Health awards DP2GM110773 and R01GM128191-02 to H.M., Research Royalty Funds from the University of Washington to H.M., the 5T32-AI055396-13 Bacterial Pathogenesis Training Grant (University of Washington) award to K.S.L., and the 1F32 AI140557-01 Ruth L. Kirschstein National Research Postdoctoral award to K.S.L. We would like to dedicate this paper to the late James Champoux, who discovered Top1 and discussed this project with us on various occasions during our time at the University of Washington.
Footnotes
SUPPLEMENTAL INFORMATION
Supplemental Information can be found online at https://doi.org/10.1016/j.celrep.2021.108797.
DECLARATION OF INTERESTS
The authors declare no competing interests.
REFERENCES
- Ashley RE, Dittmore A, McPherson SA, Turnbough CL Jr., Neuman KC, and Osheroff N (2017). Activities of gyrase and topoisomerase IV on positively supercoiled DNA. Nucleic Acids Res. 45, 9611–9624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bermejo R, Doksani Y, Capra T, Katou Y-M, Tanaka H, Shirahige K, and Foiani M (2007). Top1- and Top2-mediated topological transitions at replication forks ensure fork progression and stability and prevent DNA damage checkpoint activation. Genes Dev. 21, 1921–1936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolger AM, Lohse M, and Usadel B (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brehm SP, Staal SP, and Hoch JA (1973). Phenotypes of pleiotropic-negative sporulation mutants of Bacillus subtilis. J Bacteriol 115, 1063–1070, 10.1128/JB.115.3.1063-1070.1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brewer BJ (1988). When polymerases collide: replication and the transcriptional organization of the E. coli chromosome. Cell 53, 679–686. [DOI] [PubMed] [Google Scholar]
- Champoux JJ (2001). DNA topoisomerases: structure, function, and mechanism. Annu. Rev. Biochem 70, 369–413. [DOI] [PubMed] [Google Scholar]
- Chappidi N, Nascakova Z, Boleslavska B, Zellweger R, Isik E, Andrs M, Menon S, Dobrovolna J, Balbo Pogliano C, Matos J, et al. (2020). Fork Cleavage-Religation Cycle and Active Transcription Mediate Replication Restart after Fork Stalling at Co-transcriptional R-Loops. Mol. Cell 77, 528–541.e8. [DOI] [PubMed] [Google Scholar]
- Contreras A, and Maxwell A (1992). gyrB mutations which confer coumarin resistance also affect DNA supercoiling and ATP hydrolysis by Escherichia coli DNA gyrase. Mol. Microbiol 6, 1617–1624. [DOI] [PubMed] [Google Scholar]
- Crisona NJ, Strick TR, Bensimon D, Croquette V, and Cozzarelli NR (2000). Preferential relaxation of positively supercoiled DNA by E. coli topoisomerase IV in single-molecule and ensemble measurements. Genes Dev. 14, 2881–2892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Septenville AL, Duigou S, Boubakri H, and Michel B (2012). Replication fork reversal after replication-transcription collision. PLoS Genet. 8, e1002622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drlica K (1992). Control of bacterial DNA supercoiling. Mol. Microbiol 6, 425–433. [DOI] [PubMed] [Google Scholar]
- Drolet M, Bi X, and Liu LF (1994). Hypernegative supercoiling of the DNA template during transcription elongation in vitro. J. Biol. Chem 269, 2068–2074. [PubMed] [Google Scholar]
- Drolet M, Phoenix P, Menzel R, Massé E, Liu LF, and Crouch RJ (1995). Overexpression of RNase H partially complements the growth defect of an Escherichia coli delta topA mutant: R-loop formation is a major problem in the absence of DNA topoisomerase I. Proc. Natl. Acad. Sci. USA 92, 3526–3530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dutta D, Shatalin K, Epshtein V, Gottesman ME, and Nudler E (2011). Linking RNA polymerase backtracking to genome instability in E. coli. Cell 146, 533–543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espeli O, and Marians KJ (2004). Untangling intracellular DNA topology. Mol. Microbiol 52, 925–931. [DOI] [PubMed] [Google Scholar]
- French S (1992). Consequences of replication fork movement through transcription units in vivo. Science 258, 1362–1365. [DOI] [PubMed] [Google Scholar]
- García-Rubio M, Aguilera P, Lafuente-Barquero J, Ruiz JF, Simon M-N, Geli V, Rondón AG, and Aguilera A (2018). Yra1-bound RNA-DNA hybrids cause orientation-independent transcription-replication collisions and telomere instability. Genes Dev. 32, 965–977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gómez-González B, and Aguilera A (2019). Transcription-mediated replication hindrance: a major driver of genome instability. Genes Dev. 33, 1008–1026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffith KL, and Grossman AD (2008). Inducible protein degradation in Bacillus subtilis using heterologous peptide tags and adaptor proteins to target substrates to the protease ClpXP. Mol. Microbiol 70, 1012–1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gross CH, Parsons JD, Grossman TH, Charifson PS, Bellon S, Jernee J, Dwyer M, Chambers SP, Markland W, Botfield M, and Raybuck SA (2003). Active-site residues of Escherichia coli DNA gyrase required in coupling ATP hydrolysis to DNA supercoiling and amino acid substitutions leading to novobiocin resistance. Antimicrob. Agents Chemother 47, 1037–1046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guariglia-Oropeza V, and Helmann JD (2011). Bacillus subtilis σ(V) confers lysozyme resistance by activation of two cell wall modification pathways, peptidoglycan O-acetylation and D-alanylation of teichoic acids. J. Bacteriol 193, 6223–6232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamperl S, and Cimprich KA (2016). Conflict Resolution in the Genome: How Transcription and Replication Make It Work. Cell 167, 1455–1467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamperl S, Bocek MJ, Saldivar JC, Swigut T, and Cimprich KA (2017). Transcription-Replication Conflict Orientation Modulates R-Loop Levels and Activates Distinct DNA Damage Responses. Cell 170, 774–786.e19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardy CD, and Cozzarelli NR (2003). Alteration of Escherichia coli topoisomerase IV to novobiocin resistance. Antimicrob. Agents Chemother 47, 941–947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harwood Colin, R., and Simon M. Cutting. (1990). Molecular biological methods for Bacillus (Chichester: Wiley; ). [Google Scholar]
- Heinz S, Benner C, Spann N, Bertolino E, Lin YC, Laslo P, Cheng JX, Murre C, Singh H, and Glass CK (2010). Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol Cell 38, 576–589, 10.1016/j.molcel.2010.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hiasa H, and Marians KJ (1996). Two distinct modes of strand unlinking during theta-type DNA replication. J. Biol. Chem 271, 21529–21535. [DOI] [PubMed] [Google Scholar]
- Keszthelyi A, Minchell NE, and Baxter J (2016). The Causes and Consequences of Topological Stress during DNA Replication. Genes (Basel) 7, 134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khodursky AB, Peter BJ, Schmid MB, DeRisi J, Botstein D, Brown PO, and Cozzarelli NR (2000). Analysis of topoisomerase function in bacterial replication fork movement: use of DNA microarrays. Proc. Natl. Acad. Sci. USA 97, 9419–9424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuzminov A (2018). When DNA Topology Turns Deadly–RNA Polymerases Dig in Their R-loops to Stand Their Ground: New Positive and Negative (Super) Twists in the…. Trends Genet [DOI] [PMC free article] [PubMed]
- Lang KS, Hall AN, Merrikh CN, Ragheb M, Tabakh H, Pollock AJ, Woodward JJ, Dreifus JE, and Merrikh H (2017). Replication-Transcription Conflicts Generate R-Loops that Orchestrate Bacterial Stress Survival and Pathogenesis. Cell 170, 787–799.e18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langmead B, and Salzberg SL (2012). Fast gapped-read alignment with Bowtie 2. Nat Methods 9, 357–359, 10.1038/nmeth.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levine C, Hiasa H, and Marians KJ (1998). DNA gyrase and topoisomerase IV: biochemical activities, physiological roles during chromosome replication, and drug sensitivities. Biochim. Biophys. Acta 1400, 29–43. [DOI] [PubMed] [Google Scholar]
- Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, and Durbin R; 1000 Genome Project Data Processing Subgroup (2009). The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu LF, and Wang JC (1987). Supercoiling of the DNA template during transcription. Proc. Natl. Acad. Sci. USA 84, 7024–7027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lodge JK, Kazic T, and Berg DE (1989). Formation of supercoiling domains in plasmid pBR322. J. Bacteriol 171, 2181–2187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lynch AS, and Wang JC (1993). Anchoring of DNA to the bacterial cytoplasmic membrane through cotranscriptional synthesis of polypeptides encoding membrane proteins or proteins for export: a mechanism of plasmid hypernegative supercoiling in mutants deficient in DNA topoisomerase I. J. Bacteriol 175, 1645–1655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma J, Bai L, and Wang MD (2013). Transcription under torsion. Science 340, 1580–1583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magner DB, Blankschien MD, Lee JA, Pennington JM, Lupski JR, and Rosenberg SM (2007). RecQ promotes toxic recombination in cells lacking recombination intermediate-removal proteins. Mol. Cell 26, 273–286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mangiameli SM, Merrikh CN, Wiggins PA, and Merrikh H (2017). Transcription leads to pervasive replisome instability in bacteria. eLife 6, e19848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massé E, and Drolet M (1999). Escherichia coli DNA topoisomerase I inhibits R-loop formation by relaxing transcription-induced negative supercoiling. J. Biol. Chem 274, 16659–16664. [DOI] [PubMed] [Google Scholar]
- Maxwell A (1993). The interaction between coumarin drugs and DNA gyrase. Mol. Microbiol 9, 681–686. [DOI] [PubMed] [Google Scholar]
- Merrikh CN, and Merrikh H (2018). Gene inversion potentiates bacterial evolvability and virulence. Nat. Commun 9, 4662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merrikh H, Machón C, Grainger WH, Grossman AD, and Soultanas P (2011). Co-directional replication-transcription conflicts lead to replication restart. Nature 470, 554–557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merrikh CN, Brewer BJ, and Merrikh H (2015). The B. subtilis Accessory Helicase PcrA Facilitates DNA Replication through Transcription Units. PLoS Genet. 11, e1005289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Million-Weaver S, Samadpour AN, Moreno-Habel DA, Nugent P, Britt-nacher MJ, Weiss E, Hayden HS, Miller SI, Liachko I, and Merrikh H (2015a). An underlying mechanism for the increased mutagenesis of lagging-strand genes in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 112, E1096–E1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Million-Weaver S, Samadpour AN, and Merrikh H (2015b). Replication Restart after Replication-Transcription Conflicts Requires RecA in Bacillus subtilis. J. Bacteriol 197, 2374–2382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mirkin EV, and Mirkin SM (2005). Mechanisms of transcription-replication collisions in bacteria. Mol. Cell. Biol 25, 888–895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mostertz J, Scharf C, Hecker M, and Homuth G (2004). Transcriptome and proteome analysis of Bacillus subtilis gene expression in response to superoxide and peroxide stress. Microbiology (Reading) 150, 497–512. [DOI] [PubMed] [Google Scholar]
- Nicolas P, Mäder U, Dervyn E, Rochat T, Leduc A, Pigeonneau N, Bidnenko E, Marchadier E, Hoebeke M, Aymerich S, et al. (2012). Condition-dependent transcriptome reveals high-level regulatory architecture in Bacillus subtilis. Science 335, 1103–1106. [DOI] [PubMed] [Google Scholar]
- Nudler E (2009). RNA polymerase active center: the molecular engine of transcription. Annu. Rev. Biochem 78, 335–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nudler E (2012). RNA polymerase backtracking in gene regulation and genome instability. Cell 149, 1438–1445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohtani N, Haruki M, Morikawa M, Crouch RJ, Itaya M, and Kanaya S (1999). Identification of the genes encoding Mn2+-dependent RNase HII and Mg2+-dependent RNase HIII from Bacillus subtilis: classification of RNases H into three families. Biochemistry 38, 605–618. [DOI] [PubMed] [Google Scholar]
- Paul S, Million-Weaver S, Chattopadhyay S, Sokurenko E, and Merrikh H (2013). Accelerated gene evolution through replication-transcription conflicts. Nature 495, 512–515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng H, and Marians KJ (1993). Escherichia coli topoisomerase IV. Purification, characterization, subunit structure, and subunit interactions. J. Biol. Chem 268, 24481–24490. [PubMed] [Google Scholar]
- Pomerantz RT, and O’Donnell M (2010). Direct restart of a replication fork stalled by a head-on RNA polymerase. Science 327, 590–592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Postow L, Peter BJ, and Cozzarelli NR (1999). Knot what we thought before: the twisted story of replication. BioEssays 21, 805–808. [DOI] [PubMed] [Google Scholar]
- Postow L, Crisona NJ, Peter BJ, Hardy CD, and Cozzarelli NR (2001a). Topological challenges to DNA replication: conformations at the fork. Proc. Natl. Acad. Sci. USA 98, 8219–8226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Postow L, Ullsperger C, Keller RW, Bustamante C, Vologodskii AV, and Cozzarelli NR (2001b). Positive torsional strain causes the formation of a four-way junction at replication forks. J. Biol. Chem 276, 2790–2796. [DOI] [PubMed] [Google Scholar]
- Prado F, and Aguilera A (2005). Impairment of replication fork progression mediates RNA polII transcription-associated recombination. EMBO J. 24, 1267–1276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Promonet A, Padioleau I, Liu Y, Sanz L, Biernacka A, Schmitz A-L, Skrzypczak M, Sarrazin A, Mettling C, Rowicka M, et al. (2020). Topoisomerase 1 prevents replication stress at R-loop-enriched transcription termination sites. Nat Commun 11, 3940. 10.1038/s41467-020-17858-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramírez F, Dündar F, Diehl S, Grüning BA, and Manke T (2014). deep-Tools: a flexible platform for exploring deep-sequencing data. Nucleic Acids Res. 42, W187–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Randall JR, Hirst WG, and Simmons LA (2018). Substrate Specificity for Bacterial RNases HII and HIII Is Influenced by Metal Availability. J. Bacteriol 200, 200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson JT, Thorvaldsdóttir H, Winckler W, Guttman M, Lander ES, Getz G, and Mesirov JP (2011). Integrative genomics viewer. Nat. Bio-technol 29, 24–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samadpour AN, and Merrikh H (2018). DNA gyrase activity regulates DnaA-dependent replication initiation in Bacillus subtilis. Mol Microbiol 108, 115–127, Epub 2018 Mar 6. 10.1111/mmi.13920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanz LA, and Chédin F (2019). High-resolution, strand-specific R-loop mapping via S9.6-based DNA-RNA immunoprecipitation and high-throughput sequencing. Nat. Protoc 14, 1734–1755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schalbetter SA, Mansoubi S, Chambers AL, Downs JA, and Baxter J (2015). Fork rotation and DNA precatenation are restricted during DNA replication to prevent chromosomal instability. Proc. Natl. Acad. Sci. USA 112, E4565–E4570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smits WK, Goranov AI, and Grossman AD (2010). Ordered association of helicase loader proteins with the Bacillus subtilis origin of replication in vivo. Mol. Microbiol 75, 452–461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smits WK, Merrikh H, Bonilla CY, and Grossman AD (2011). Primosomal proteins DnaD and DnaB are recruited to chromosomal regions bound by DnaA in Bacillus subtilis. J. Bacteriol 193, 640–648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stolz R, Sulthana S, Hartono SR, Malig M, Benham CJ, and Chedin F (2019). Interplay between DNA sequence and negative superhelicity drives R-loop structures. Proc. Natl. Acad. Sci. USA 116, 6260–6269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugino A, Higgins NP, Brown PO, Peebles CL, and Cozzarelli NR (1978). Energy coupling in DNA gyrase and the mechanism of action of novobiocin. Proc. Natl. Acad. Sci. USA 75, 4838–4842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tadesse S, and Graumann PL (2006). Differential and dynamic localization of topoisomerases in Bacillus subtilis. J. Bacteriol 188, 3002–3011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuduri S, Crabbé L, Conti C, Tourrière H, Holtgreve-Grez H, Jauch A, Pantesco V, De Vos J, Thomas A, Theillet C, et al. (2009). Topoisomerase I suppresses genomic instability by preventing interference between replication and transcription. Nat. Cell Biol 11, 1315–1324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vos SM, Tretter EM, Schmidt BH, and Berger JM (2011). All tangled up: how cells direct, manage and exploit topoisomerase function. Nat. Rev. Mol. Cell Biol 12, 827–841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang JC (2002). Cellular roles of DNA topoisomerases: a molecular perspective. Nat. Rev. Mol. Cell Biol 3, 430–440. [DOI] [PubMed] [Google Scholar]
- Wang JD, Berkmen MB, and Grossman AD (2007). Genome-wide coorientation of replication and transcription reduces adverse effects on replication in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 104, 5608–5613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu HY, Shyy SH, Wang JC, and Liu LF (1988). Transcription generates positively and negatively supercoiled domains in the template. Cell 53, 433–440. [DOI] [PubMed] [Google Scholar]
- Zechiedrich EL, and Cozzarelli NR (1995). Roles of topoisomerase IV and DNA gyrase in DNA unlinking during replication in Escherichia coli. Genes Dev. 9, 2859–2869. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Datasets generated during this study are available from NCBI SRA project ID PRJNA691533.