
Figure 1.
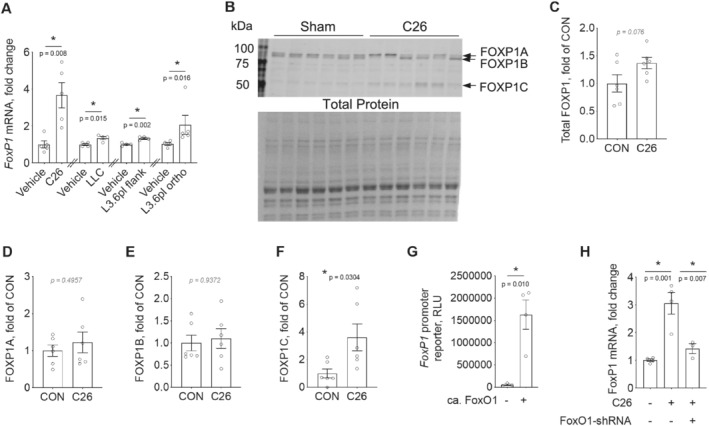
FoxP1 is up‐regulated in multiple models of cancer cachexia in a FoxO1‐dependent manner. (A) Changes in FoxP1 mRNA in mouse skeletal muscle in various experimental models of cancer cachexia, including the subcutaneous murine colon 26 adenocarcinoma (C26) and Lewis lung carcinoma (LLC) models, and the human pancreas‐liver (L3.6pl) xenograft models, in which human L3.6pl cells were injected into the flank or orthotopically (ortho) into the pancreas. Note that independent unpaired two‐tailed t‐tests were performed for the different cachexia models. (B–F) FOXP1 protein expression in skeletal muscle of C26 tumour‐bearing and non‐tumour‐bearing (Sham) mice. In panel (C), total FOXP1 expression was quantified by summing the signal from FOXP1A, FOXP1B, and FOXP1C and normalizing to total protein. (G) Transfection of rat solei with a constitutively active FoxO1 expression plasmid (ca. FoxO1) is sufficient to increase FoxP1 transcription, in vivo, as highlighted by its ability to increase a luciferase reporter gene driven by the FoxP1 promoter (RLU = relative light unit). (H) FoxO1 knockdown in mouse tibialis anterior, through transduction of muscles with AAV9 vector encoding FoxO1‐shRNA (or scrambled‐shRNA), prevents the C26‐induced upregulation of FoxP1 mRNA. For panels (A)–(G), depending on data distribution, unpaired two‐tailed t‐tests or Mann–Whitney tests were performed to test for statistical differences between groups. A one‐way analysis of variance (ANOVA) was conducted to test for statistical differences between means in panel (H). Data are reported as mean ± SEM.