

ABSTRACT
Children have elevated fever risk 1 to 2 weeks after the first dose of a measles-containing vaccine (MCV), which is likely affected by genetic, immunologic, and clinical factors. Fever after MCV is associated with febrile seizures, though may also be associated with higher measles antibody titers. This exploratory study investigated genetic and immunologic associations with a fever after MCV. Concurrent with a randomized Phase 3 clinical trial of 12–15-month-olds who received their first measles-mumps-rubella (MMR) vaccine in which parents recorded post-vaccination temperatures daily, we consented a subset to collect additional blood and performed human leukocyte antigens (HLA) typing. Association between fever 5–12 days after MMR (“MMR-associated”) and HLA type was assessed using logistic regression. We compared 42-day post-vaccination geometric mean titers (GMT) to measles between children who did and did not have fever using a t-test. We enrolled 86 children and performed HLA typing on 82; 13 (15.1%) had MMR-associated fever. Logistic regressions identified associations between MMR-associated fever and HLA Class I loci A-29:02 (P = .036), B-57:01 (P = .018), C-06:02 (P = .006), C-14:02 (P = .022), and Class II loci DRB1-15 (P = .045). However, Bonferroni's adjustment for multiple comparisons suggests that these associations could have been due to chance. Ninety-eight percent of children had protective antibody titers to measles; however, GMT was higher among those with fever compared with children without fever (P = .006). Fever after the measles vaccine correlated with genetic factors and higher immune response. This study suggests a possible genetic susceptibility to MMR-associated fever.
KEYWORDS: MMR, MMRV, vaccine, genetic factors, fever, antibody, titers
Introduction
Fever is a common nonspecific immune response to infection and immunization and may have a beneficial role in response to infection.1,2 However, in the context of immunization, fever is considered an adverse event that leads to increased medical visits.3 Parental concerns about vaccine adverse events, even mild ones such as fever, have been associated with parental reluctance to vaccinate.4,5 Parental vaccine hesitancy negatively impacts public health,6 as evidenced by 2018–19 measles outbreak in the United States which saw the largest number of cases since 1992 and was mainly attributable to under vaccination.7 Elucidating a positive role for fever after vaccination could reassure parents and may help address vaccine hesitancy.
Several lines of evidence suggest that genetic factors influence susceptibility to fever following the first dose of MCV. MCV predictably causes fever 7–10 days after vaccination in 5%–15% of the vaccinees8–16 and this fever is strongly associated with febrile seizures 7–10 days after MCV.8,13,15 A large Danish genome-wide association study demonstrated that susceptibility to measles, mumps, rubella (MMR) vaccine-associated febrile seizures have a genetic basis.17 A Vaccine Safety Datalink (VSD) study evaluating 946,806 children found that children with siblings who had fever 7–10 days after a MCV was more than 3 times as likely themselves to have fever 7–10 days after a MCV.18 More recently, we reported that parental history of fever and fever with respiratory infection was also associated with an increased risk of fever in their child 7–10 days after MCV.19
Fever after MMR may be related to higher immune response,20 and genetics influence the immune response to MMR vaccine. An analysis of 5 MCV clinical trials detected an association between post-vaccination fever and higher immune responses, with the level of post-vaccination measles antibody titers positively associated with fever.20 The heritability of antibody titers in response to measles vaccination varies between 49% and 88.5%.21–23 Studies have reported associations between differential immune responses to MMR and human leukocyte antigen (HLA) alleles,24–27 as well as with other cellular receptors that influence measles vaccine responses.28–36
Together these studies provide support for the hypothesis that genetic and biological factors influence susceptibility to fever following measles vaccination. Analyzing biospecimens previously collected from one-year-olds enrolled in a randomized-controlled MMR vaccine clinical trial,16 the objectives of this exploratory study were to investigate the association between HLA type and fever after MCV and between fever after MCV and antibody titers.
Materials and methods
Study design and population
This study was conducted by separately enrolling subjects who had already been enrolled in a large-randomized, controlled Phase 3 clinical trial evaluating an investigational MMR vaccine (licensed in >100 countries). The Phase 3 trial enrolled subjects aged 12–15 months to receive either a single dose of an investigational MMR vaccine or of the US licensed MMR vaccine (MMR II, Merck & Co) as described.16 Subjects had blood samples collected before and 42 days after vaccination. As part of the clinical trial, parents were given thermometers, taught how to measure daily temperatures, and instructed to record the highest temperature each day on a diary card prospectively for 42 days after vaccination. The clinical trial enrolled a total of 1736 subjects globally. The incidence of fever 5–12 days post-vaccination was comparable in the clinical trial, with similar rates of fever and other MMR-specific symptoms reported after both MMR vaccines.16 Kaiser Permanente Northern California (KPNC), as a participating clinical trial site, enrolled 341 of the 1736 subjects.
Concurrent with the per-protocol activities for the above clinical trial, KPNC separately consented the parents/legally authorized representatives of enrolled children to obtain and store additional vials of blood after collecting the above per-protocol blood samples. Due to resource and time constraints, only a portion of the 341 children were enrolled in this separate study. The goal of this separate study was to investigate genetic and biologic factors related to developing fever associated with the MMR vaccine using stored biospecimens. This current report focuses on the analyses of these stored biospecimens.
The KPNC and Stanford University Institutional Review Boards approved this study.
Biospecimen collection and storage
We obtained approximately 6 ml of blood in an EDTA tube, inverted to mix 8–10 times, and immediately spun the tube for 10 min at 1,000–3000 G’s. We removed the plasma and the buffy coat layers placed each into separate cryovials which were immediately frozen at −20°C, where they remained for several months. The plasma and buffy coat cryovials were subsequently transferred and stored frozen at −80°C, where they remained until they were processed and analyzed.
DNA extraction
DNA from buffy coat specimens were extracted using the Qiagen DNA extraction kit (catalog number 56504) following standard protocol. DNA samples underwent quality control in three ways: NanoDrop, Qubit, and fragment analyzer. Nanodrop readings were used to assess the overall sample quality. Qubit reading was used for quantification (Thermofisher catalog number Q32854) following standard protocol. Fragment analyzer traces were used to assess sample quality following standard protocol. All samples met the standard quality for library synthesis.
HLA library preparation and sequencing
We used 400 ng of intact and not fragmented genomic DNA to generate HLA libraries using Illumina TruSight HLAv2 library prep kit (Catalog # 20000215) following standard protocol. The TruSight HLA v2 sequencing panel covers all commonly typed HLA loci. A single assay results in the sequencing of 11 HLA loci, 3 of which were Class I (HLA A, HLA B and HLA C) and 8 were Class II (DRB1, DRB3, DRB4, DRB5, DPA1, DPB1, DQA1, and DQB1). All the sequencing was done on MiSeq using a 2 × 150 pair end kit following standard protocol.
Antibody titers
Measles-specific immunoglobulin G (IgG) antibodies (mIU/mL) were measured using commercial ELISA kits as described16 and were provided by the sponsor of the Phase 3 trial. Measles titers were considered protective if the post-vaccination antibody concentration was ≥200 mIU/mL for children who were seronegative before vaccination (defined as ≤150 mIU/mL) as described.16
Statistical analyses
For all analyses, to be consistent with the original clinical trial,16 we considered a temperature ≥100.4°F(38°C) between days 5 and 12 recorded on the daily diary cards as a fever, henceforth known as “MMR-associated fever.”
We described the distribution of selected demographic factors (Table 1) and the proportion of each HLA type among children who did and did not have MMR-associated fever. We assessed genotypes according to a dominant inheritance pattern. Participants were classified as a carrier of the allele if at least one allele copy of the allele of interest was present. We determined allele frequencies across diverse populations in the US using the US National Marrow Donor Program report.37 This report presents allele frequencies across population subgroups, including African American, Asian, European American, and Hispanic cohorts in the US. Based on these reported allele frequencies, we calculated the expected proportion of individuals in each subpopulation carrying at least one copy of the allele of interest (carriers) by assuming Hardy-Weinberg Equilibrium and taking the square of the allele frequency (q^2) and adding it to 2*alleleFreq*(1-alleleFreq).38 We defined the expected population carrier frequency range for each allele as the range of the proportion of individuals who would be expected to carry at least one of these alleles across diverse US subpopulations, where the low end of the range reflects the calculated carrier frequency based on the subpopulation where the allele was reported to be least common and where the high end of the range reflects the calculated carrier frequency based on the subpopulation where the allele was reported to be most common. Thus, we expect the carrier frequency in our cohort to fall within that range regardless of the distribution of specific genetic ancestries. We assessed for an association between MMR-associated fever and HLA type using unadjusted logistic regression. We only report HLA genes where ≥50% of the children had two typed alleles.
Table 1.
Racial and ethnic demographics among children with and without MMR-associated fever
| Children without MMR-Associated Fevera N = 66 (%) |
Children with MMR-Associated Feverb N = 13 (%) | |
|---|---|---|
| Race | ||
| White – Caucasian/European Heritage | 32 (48.5) | 6 (46.2) |
| African Heritage/African American | 7 (10.6) | 0 |
| Asian | 14 (21.2) | 4 (30.8) |
| American Indian or Alaskan Native | 3 (4.5) | 2 (15.4) |
| Multiple Races | 10 (15.2) | 1 (7.7) |
| Ethnicity | ||
| Non-Hispanic | 58 (87.8) | 10 (76.9) |
| Hispanic | 8 (12.2) | 3 (23.1) |
aDoes not include 7 subjects for whom fever data was missing (3 White, 2 Asian and 2 African Heritage).
b14.5% had fever after a single dose of an investigational MMR and 16.1% after the US-licensed MMR vaccine.
For the antibody titer analyses, we compared day 42 post-vaccination measles-specific geometric mean titers (GMT) between children who did and did not have MMR-associated fever using a t-test after natural logarithm transformation to achieve normality. Box plots of geometric means were created using the Surveymeans procedure in SAS.
Analyses were performed using SAS, version 9.4 (SAS Institute, Inc., Cary, North Carolina), and R v3.1.2 (R Core Team, 2012) glm package.
P values were two-sided and those ≤0.05 were considered as statistically significant. We adjusted the P values for the multiplicity of hypotheses tested using Bonferroni correction. We regarded the HLA results as exploratory given the small sample size.
Results
We separately consented and enrolled 86 children and, of these, were able to obtain buffy coat specimens on 83/86 (97%). One child subsequently withdrew consent just for HLA typing, for a final of 82 children on whom we performed HLA typing. Among the total 86 children, 13 (15.1%) had MMR-associated fever (14.5% after investigational MMR and 16.1% after MMR II). Racial and ethnic demographics were broadly similar between the fever and no fever groups, except that none of the children with fever was of Black race (regardless of ethnicity) versus 10.6% of those without fever (Table 1). Comparisons were limited due to the small numbers of children.
HLA sequence data were assessed for a total of 187 alleles. It was available for both alleles for 94% of the children for HLA A, 95% for HLA B, 87% for HLA C, 100% for HLA DPA1, 77% for DPB1, 99% for HLA DQA1, 94% HLA DQB1, and 98% for HLA DRB1. We excluded HLA DRB3, DRB4, and DRB5 alleles because <50% of the children had two typed alleles.
Logistic regressions identified associations between MMR-associated fever and 4 HLA Class I loci and 1 Class II loci, with 1 additional Class II loci having a borderline association (Table 2). None of the children with MMR-associated fever were homozygous for these five loci. These five loci occurred more commonly among children with MMR-associated fever than would be expected based on allele frequencies across diverse US populations. For example, we expected that 3%–7% of our study population would have HLA A-29; however, we observed that 25% of the children with MMR-associated fever had HLA A-29, while only 5% of those without fever had HLA A-29 (P = .036). In contrast, HLA C-4 was expected to occur in 15%–33% of the study population and we noted that 27.1% of the children without fever and 8.3% of the children with MMR-associated fever had the HLA C-4 loci (P = .189; Supplemental Table). However, adjusting for the multiplicity of hypotheses tested (n = 71) using the Bonferroni correction suggests that these associations could be due to chance. Out of the 187 total alleles, there were 66 alleles that were identified in at least two children, with at least one of the two being in the group with MMR-associated fever. These 66 were not associated with MMR-associated fever (Supplemental Table).
Table 2.
Association between human leukocyte antigen allele and MMR-associated fever
| Allele Associated with Fever | Allele frequency across diverse US subpopulationsa (%) | Expected proportion with at least one allele (%) | Proportion among children without MMR-associated Fever (%)b | Proportion among children with MMR-associated Fever (%) | Odds Ratioc (95% Confidence Interval) | P value | P value after Bonferroni correction |
|---|---|---|---|---|---|---|---|
| HLA Class I | |||||||
| A-29:02 | 1–4 | 2–7 | 3/60 (5) | 3/12 (25) | 6.3 (1.1, 36.4) | 0.036 | 1 |
| B-57:01 | 0–4 | 0–7 | 2/63 (3.2) | 3/12 (25) | 10.2 (1.5, 69.4) | 0.018 | 1 |
| C-06:02 | 5–9 | 10–17 | 2/55 (3.6) | 4/12 (33.3) | 13.3 (2.1, 84.5) | 0.006 | 0.426 |
| C-14:02 | 1–5 | 2–10 | 0/65 (0) | 2/12 (16.7) | NE (1.63, NE)d | 0.022 | 1 |
| HLA Class II | |||||||
| DRB1-04:01 | 1–9 | 2–17 | 4/65 (6.2) | 3/12 (25) | 5.1 (1.0, 26.5) | 0.054 | 1 |
| DRB1-15 | 2–15 | 4–27 | 18/65 (27.7) | 7/12 (58.3) | 3.7 (1.0, 13.0) | 0.045 | 0.426 |
aWe calculated the expected carrier frequency from allele frequencies37 assuming Hardy-Weinberg Equilibrium.38
bDenominator includes children for whom both HLA type and fever data were available.
cComparing children with MMR-associated fever vs those without MMR-associated fever.
dThe lower bound CI was based on the conditional maximum likelihood estimate of the odds ratio. P value was calculated as an exact P value.
Measles antibody titer data were available for 84 children, of whom 82 (98%) had protective titers to measles 42 days after vaccination. Among the children for whom we had both titer and fever data (n = 77), post-vaccination GMT was higher among those with MMR-associated fever (4609, 95% confidence interval [CI] 3629, 5853) than among children who did not have a fever (2918, 95% CI 2318, 3673, P = .006; Figure 1). Higher post-vaccination GMT and MMR-associated fever were also noted among the 341 children enrolled in the trial at KPNC (GMT among children with fever: 4022 [95% CI 3209, 5042] vs children with no fever: 3082 [95% CI 2792, 3402]; P = .032).
Figure 1.
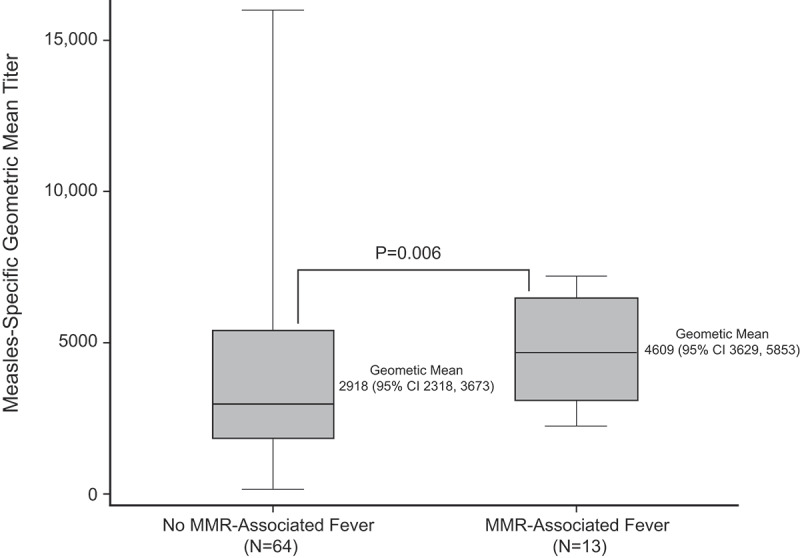
Geometric mean titers (GMT) against measles among children with and without fever after MMR
The horizontal line within each box denotes the median GMT for each group.
Discussion
This study investigated whether genetic factors influence susceptibility to fever and if fever is associated with increased immunogenicity to MCV. In this exploratory study which used biospecimens previously collected during a randomized-controlled MMR vaccine clinical trial, we identified 5 HLA alleles associated with a fever after the MMR vaccine. However, after adjusting for the multiplicity of testing these findings could be due to chance alone because of the small sample size. While the current exploratory study does not allow us to draw firm conclusions regarding specific associations between fever after MMR and HLA types due to its small size, and a stringent Bonferroni correction, it does suggest based on effect size with confidence intervals excluding the null value that specific genetic factors correlate with a fever after MCV. Importantly, our study provides additional support for the growing body of evidence that genetics influences susceptibility to fever after the MMR vaccine.
Genetic factors such as HLA type have not previously been directly correlated with a fever after MCV. Although our findings must be interpreted with caution due to the small size of this study, the HLA alleles identified here as potentially being correlated with susceptibility to fever after the MMR vaccine can serve as important targets in future studies. To put our findings in the context of the US population, we also calculated the expected population carrier frequencies to reflect the range of allele frequencies (from least to most common) that have been reported in populations across racially diverse cohorts in the US. These ranges help interpretation of our findings by providing context as to how our study population compared with alleles which are generally more or less common across diverse communities in the US. For example, based on prior studies reflecting many subpopulations, we expected just 3%–7% of our participants to have the HLA A-29 allele.37 In other words, in US populations where the HLA A-29 allele is reported to be most common, we expect to find just 7% of the people with that allele. However, we found that allele in 25% of our participants who experienced fever, which is substantially higher than in the population with the highest reported allele frequency.
Several candidate HLA alleles we identified in association with a fever after the MMR vaccine have been associated previously with a viral response. For example, HLA-B 57:01 has previously been reported to be strongly associated with a hypersensitivity syndrome characterized by fever, malaise, and gastrointestinal symptoms in up to 8% of those starting the guanosine analog antiviral abacavir.39–42 Another potential HLA allele that may be worthy of future investigations includes HLA-C 06:02 which was seen more than expected among children with fever (33% vs 10%–17% expected). Finally, although the association between HLA-DRB1-04:01 and fever was borderline, HLA-DRB1-04:05 has been associated with resistance to enteric fever.43
These data implicating genes involved with fever are consistent with prior studies on heritability of fever following the measles vaccine. Studies showing a role of genetics in fever response include a VSD study reporting that children with siblings who had fever 7–10 days after a MCV was more than 3 times more likely themselves to have fever 7–10 days after a MCV.18 Furthermore, a recent study that parental history of fever and fever with respiratory infection were associated with fever in their child 7–10 days after MCV.19
We directly demonstrated an association between fever after MMR and higher measles titers, providing an important link between a stronger immune response to MMR vaccine and fever after the MMR vaccine. Our results are consistent with a meta-analysis which assessed pre-licensure measles, mumps, rubella, varicella vaccine clinical trials, and reported an association between post-vaccination fever and higher immune responses, with the level of post-vaccination measles antibody titers positively associated with fever.20 Studies showing lower antibody responses in infants given antipyretics prior to vaccination also lend support to the fever pathway aiding in the immunogenicity of vaccines.44 Importantly, a lack of MMR-associated fever did not indicate inadequate immune response to MMR as nearly all subjects, regardless of whether they had a fever, were seroprotected against measles. Rather, genetic variation affecting fever after MMR may reflect an improved immune response to measles and suggests that beneficial immune effects may be initiated through the fever pathways.2
The association between fever after MMR and higher measles titers strengthens the evidence that genetics impact susceptibility to fever after the MMR vaccine. Differential immune responses to the MMR vaccine has been reported in associations with genetic variation in the immune system,21–23 including HLA alleles,24–26 cytokine and cytokine receptors,28 viral pattern recognition receptors which influence measles vaccine responses,30–36 and measles cellular receptors (CD46, SLAM, and others).29 A recent large population-based study evaluated measles titers among adolescents and adults 11–41 years of age previously vaccinated at differing times with MMR (presumably most were second MMR doses) identified HLA alleles of B 57:01, DRB1-15:05, and DQB1-06:02 as being associated with circulating antibody titers to MCV.27 This study suggested associations between DQB1*06:02 and DRB1*15:05 with higher measles antibody titers and between B*57:01 alleles with lower titers. Important differences between this report and our current study was that we only included toddlers receiving their first dose of MMR vaccine thereby focusing exclusively on the primary antibody response to MMR. It is possible that genetic factors which may be associated with the primary measles vaccine immune response may differ from those associated with the maintenance of circulating antibodies to the measles vaccine.
Specific genes implicated with MMR-associated febrile seizures in a prior GWAS have also been associated with immunity to the measles vaccine. In particular, variants IFI44L and CD46 involved with MMR-associated febrile seizures17 have also been associated with variability in measles antibody titers following vaccination in another GWAS.45 Notably, the CD46 protein is the cell-surface receptor for the measles virus, and variation in its expression has been associated with variation in measles-specific IgG antibody levels as well as with IL-6, TNFα, and IFNα responses following measles vaccine.29 The IFI44L protein belongs to the interferon-stimulated genes and is likely involved in innate antiviral immunity.46 Based on prior GWAS, genetic risk factors for vaccine-related febrile seizures seem distinct from febrile seizures generally and appear driven by the immune response.17
Strengths of this study were that we collected biospecimens during a randomized-controlled clinical trial which ensured that all children were between 12 and 15 months of age, were all receiving their first dose of MMR, all antibody titer data were obtained 42 days post-vaccination, and importantly, all post-vaccination fever data were prospectively recorded. Our study included data on genetic, fever, and antibody titer level and we were able to analyze all three factors in the same population.
Our study had notable limitations, the most important of which was its small size. For this reason, we consider the identified HLA alleles as preliminary and need confirmation in a large study. Although the subjects were enrolled as part of a sub-study of the larger randomized clinical trial and reflected the racial distribution of all subjects enrolled within KPNC, it is likely they did not reflect the racial and ethnic background of all subjects who were enrolled in the larger global clinical trial. For example, if an HLA allele was more common in a racial subgroup that also had more fever, we may have identified that the HLA allele as being associated with fever due to the effects of confounding from population structure because of our sample size did not allow for adjustment for race/ethnicity. In addition, we were limited in our ability to assess several HLA alleles (DRB3, DRB4, and DRB5) because these assays did not perform well.
In conclusion, utilizing MCV-associated fever and febrile seizures as a model we can begin to understand the link between population health (surveillance of a large population for vaccine adverse events) and personalized medicine (the genetic basis for susceptibility to MCV-associated fever and febrile seizures), and the implications that these findings might have on the precision vaccinology field. Vaccine-associated fever, potentially due in part to genetic variation, may serve as a proxy for an immunologically meaningful phenotype and may serve to reassure parents and benefit public health by linking it to improved effectiveness.
Supplementary Material
Acknowledgments
We would like to thank Charlie Chao, MA, CCRP for assistance with sample preparation, shipping, and data management.
Funding Statement
This work was supported by the U.S. Department of Health and Human Services, under a cooperative agreement with the National Vaccine Program Office, to NPK. OZ was funded in part by grant 1K01AI139275-01 from the National Institute of Allergy and Infectious Diseases.
Disclosure of potential conflicts of interest
NPK report research support from GlaxoSmithKline for the Phase 3 clinical trial from which the patients in the study were recruited. The measles vaccines which are the focus of this study were provided by GlaxoSmithKline as part of this clinical trial. NPK has received research grants from GlaxoSmithKline, Sanofi Pasteur, Merck, Pfizer, and Protein Science. All other authors have no conflicts of interest relevant to this article to disclose.
All authors attest they meet the ICMJE criteria for authorship.
Supplementary data
Supplemental data for this article can be accessed on the publisher’s website
References
- 1.Launey Y, Nesseler N, Malledant Y, Seguin P.. Clinical review: fever in septic ICU patients–friend or foe? Crit Care. 2011;15:222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Evans SS, Repasky EA, Fisher DT. Fever and the thermal regulation of immunity: the immune system feels the heat. Nat Rev Immunol. 2015;15:335–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wilson K, Hawken S, Kwong JC, Deeks S, Crowcroft NS, Van Walraven C, Potter BK, Chakraborty P, Keelan J, Pluscauskas M, et al. Adverse events following 12 and 18 month vaccinations: a population-based, self-controlled case series analysis. PLoS One. 2011;6:e27897. doi: 10.1371/journal.pone.0027897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Saada A, Lieu TA, Morain SR, Zikmund-Fisher BJ, Wittenberg E. Parents’ choices and rationales for alternative vaccination schedules: a qualitative study. Clin Pediatr (Phila). 2014;54(3):236–43. [DOI] [PubMed] [Google Scholar]
- 5.Williams SE. What are the factors that contribute to parental vaccine-hesitancy and what can we do about it? Hum Vaccin Immunother. 2014;10:2584–96. doi: 10.4161/hv.28596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zipprich J, Winter K, Hacker J, Xia D, Watt J, Harriman K. Measles outbreak - california, december 2014-february 2015. MMWR Morb Mortal Wkly Rep. 2015;64:153–54. [PMC free article] [PubMed] [Google Scholar]
- 7. McDonald R, Ruppert PS, Souto M, Johns DE, McKay K, Bessette N, McNulty LX, Crawford JE, Bryant P, Mosquera MC, et al. Notes from the field: measles outbreaks from imported cases in orthodox Jewish communities - New York and New Jersey, 2018–2019. MMWR Morb Mortal Wkly Rep. 2019;68:444–45. doi: 10.15585/mmwr.mm6819a4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Klein NP, Lewis E, Fireman B, Hambidge SJ, Naleway A, Nelson JC, Belongia EA, Yih WK, Nordin JD, Hechter RC, et al. Safety of measles-containing vaccines in 1-year-old children. Pediatrics. 2015;135:e321–9. doi: 10.1542/peds.2014-1822. [DOI] [PubMed] [Google Scholar]
- 9.Strebel PM, Papania MJ, Dayan GH, Halsey NA. Measles Vaccine. In: Plotkin SA, Orenstein W, Offit PA, editors. Vaccines. (Basel): Saunders Elsevier; 2008. p. 353–98. [Google Scholar]
- 10.Hirayama M. Measles vaccines used in Japan. Rev Infect Dis. 1983;5:495–503. doi: 10.1093/clinids/5.3.495. [DOI] [PubMed] [Google Scholar]
- 11.Peltola H, Heinonen OP. Frequency of true adverse reactions to measles-mumps-rubella vaccine. A double-blind placebo-controlled trial in twins. Lancet. 1986;1:939–42. [DOI] [PubMed] [Google Scholar]
- 12.Blatter MM, Klein NP, Shepard JS, Leonardi M, Shapiro S, Schear M, Mufson MA, Martin JM, Varman M, Grogg S, et al. Immunogenicity and safety of two tetravalent (measles, mumps, rubella, varicella) vaccines coadministered with hepatitis a and pneumococcal conjugate vaccines to children twelve to fourteen months of age. Pediatr Infect Dis J. 2012;31:e133–40. doi: 10.1097/INF.0b013e318259fc8a. [DOI] [PubMed] [Google Scholar]
- 13.Klein NP, Fireman B, Yih WK, Lewis E, Kulldorff M, Ray P, Baxter R, Hambidge S, Nordin J, Naleway A, et al. Measles-mumps-rubella-varicella combination vaccine and the risk of febrile seizures. Pediatrics. 2010;126:e1–8. doi: 10.1542/peds.2010-0665. [DOI] [PubMed] [Google Scholar]
- 14.Klein NP, Shepard J, Bedell L, Odrljin T, Dull P. Immunogenicity and safety of a quadrivalent meningococcal conjugate vaccine administered concomitantly with measles, mumps, rubella, varicella vaccine in healthy toddlers. Vaccine. 2012;30:3929–36. doi: 10.1016/j.vaccine.2012.03.080. [DOI] [PubMed] [Google Scholar]
- 15.Rowhani-Rahbar A, Fireman B, Lewis E, Nordin J, Naleway A, Jacobsen SJ, Jackson LA, Tse A, Belongia EA, Hambidge SJ, et al. Effect of age on the risk of Fever and seizures following immunization with measles-containing vaccines in children. JAMA Pediatr. 2013;167:1111–17. doi: 10.1001/jamapediatrics.2013.2745. [DOI] [PubMed] [Google Scholar]
- 16.Group MS. Safety and immunogenicity of an upper-range release titer measles-mumps-rubella vaccine in children vaccinated at 12 to 15 months of age: a phase III, randomized study. Hum Vaccin Immunother. 2018;14:2921–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Feenstra B, Pasternak B, Geller F, Carstensen L, Wang T, Huang F, Eitson JL, Hollegaard MV, Svanström H, Vestergaard M, et al. Common variants associated with general and MMR vaccine-related febrile seizures. Nat Genet. 2014;46:1274–82. doi: 10.1038/ng.3129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Klein NP, Lewis E, McDonald J, Fireman B, Naleway A, Glanz J, Jackson LA, Donahue JG, Jacobsen SJ, Weintraub E, et al. Risk factors and familial clustering for fever 7–10days after the first dose of measles vaccines. Vaccine. 2017;35:1615–21. doi: 10.1016/j.vaccine.2017.02.013. [DOI] [PubMed] [Google Scholar]
- 19.Zerbo O, Glanternik JR, Modaressi S, Goddard K, Ross P, Lewis N, Klein NP. Parental risk factors for fever in their children 7–10 days after the first dose of measles-containing vaccines. Hum Vaccin Immunother. 2019;16(4):875–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kuter BJ, Brown ML, Hartzel J, Williams WR, EvesiKaren A, Black S, Shinefield H, Reisinger KS, Marchant CD, Sullivan BJ, et al. Safety and immunogenicity of a combination measles, mumps, rubella and varicella vaccine (ProQuad). Hum Vaccin. 2006;2:205–14. doi: 10.4161/hv.2.5.3246. [DOI] [PubMed] [Google Scholar]
- 21.Brunell PA, Weigle K, Murphy MD, Shehab Z, Cobb E. Antibody response following measles-mumps-rubella vaccine under conditions of customary use. JAMA. 1983;250:1409–12. doi: 10.1001/jama.1983.03340110023025. [DOI] [PubMed] [Google Scholar]
- 22.Tan PL, Jacobson RM, Poland GA, Jacobsen SJ, Pankratz VS. Twin studies of immunogenicity–determining the genetic contribution to vaccine failure. Vaccine. 2001;19:2434–39. doi: 10.1016/S0264-410X(00)00468-0. [DOI] [PubMed] [Google Scholar]
- 23.Schaid DJ, Haralambieva IH, Larrabee BR, Ovsyannikova IG, Kennedy RB, Poland GA. Heritability of vaccine-induced measles neutralizing antibody titers. Vaccine. 2017;35:1390–94. doi: 10.1016/j.vaccine.2017.01.078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ovsyannikova IG, Pankratz VS, Vierkant RA, Jacobson RM, Poland GA. Human leukocyte antigen haplotypes in the genetic control of immune response to measles-mumps-rubella vaccine. J Infect Dis. 2006;193:655–63. doi: 10.1086/500144. [DOI] [PubMed] [Google Scholar]
- 25.Ovsyannikova IG, Jacobson RM, Vierkant RA, Pankratz VS, Poland GA. HLA supertypes and immune responses to measles-mumps-rubella viral vaccine: findings and implications for vaccine design. Vaccine. 2007;25:3090–100. doi: 10.1016/j.vaccine.2007.01.020. [DOI] [PubMed] [Google Scholar]
- 26.Ovsyannikova IG, Pankratz VS, Vierkant RA, Jacobson RM, Poland GA. Consistency of HLA associations between two independent measles vaccine cohorts: a replication study. Vaccine. 2012;30:2146–52. doi: 10.1016/j.vaccine.2012.01.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ovsyannikova IG, Schaid DJ, Larrabee BR, Haralambieva IH, Kennedy RB, Poland GA. A large population-based association study between HLA and KIR genotypes and measles vaccine antibody responses. PLoS One. 2017;12:e0171261. doi: 10.1371/journal.pone.0171261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Haralambieva IH, Ovsyannikova IG, Pankratz VS, Kennedy RB, Jacobson RM, Poland GA. The genetic basis for interindividual immune response variation to measles vaccine: new understanding and new vaccine approaches. Expert Rev Vaccines. 2013;12:57–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ovsyannikova IG, Haralambieva IH, Vierkant RA, O’Byrne MM, Jacobson RM, Poland GA. The association of CD46, SLAM and CD209 cellular receptor gene SNPs with variations in measles vaccine-induced immune responses: a replication study and examination of novel polymorphisms. Hum Hered. 2011;72:206–23. doi: 10.1159/000331585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dhiman N, Poland GA, Cunningham JM, Jacobson RM, Ovsyannikova IG, Vierkant RA, Wu Y, Pankratz VS. Variations in measles vaccine-specific humoral immunity by polymorphisms in SLAM and CD46 measles virus receptors. J Allergy Clin Immunol. 2007;120:666–72. doi: 10.1016/j.jaci.2007.04.036. [DOI] [PubMed] [Google Scholar]
- 31.Dhiman N, Ovsyannikova IG, Vierkant RA, Ryan JE, Pankratz VS, Jacobson RM, Poland GA. Associations between SNPs in toll-like receptors and related intracellular signaling molecules and immune responses to measles vaccine: preliminary results. Vaccine. 2008;26:1731–36. doi: 10.1016/j.vaccine.2008.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Clifford HD, Richmond P, Khoo SK, Zhang G, Yerkovich ST, Le Souef PN, Hayden CM. SLAM and DC-SIGN measles receptor polymorphisms and their impact on antibody and cytokine responses to measles vaccine. Vaccine. 2011;29:5407–13. doi: 10.1016/j.vaccine.2011.05.068. [DOI] [PubMed] [Google Scholar]
- 33.Clifford HD, Hayden CM, Khoo SK, Zhang G, Le Souef PN, Richmond P. CD46 measles virus receptor polymorphisms influence receptor protein expression and primary measles vaccine responses in naive Australian children. Clin Vaccine Immunol. 2012;19:704–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Clifford HD, Yerkovich ST, Khoo SK, Zhang G, Upham J, Le Souef PN, Richmond P, Hayden CM. Toll-like receptor 7 and 8 polymorphisms: associations with functional effects and cellular and antibody responses to measles virus and vaccine. Immunogenetics. 2012;64:219–28. doi: 10.1007/s00251-011-0574-0. [DOI] [PubMed] [Google Scholar]
- 35.Clifford HD, Yerkovich ST, Khoo SK, Zhang G, Upham J, Le Souef PN, Richmond P, Hayden CM. TLR3 and RIG-I gene variants: associations with functional effects on receptor expression and responses to measles virus and vaccine in vaccinated infants. Hum Immunol. 2012;73:677–85. doi: 10.1016/j.humimm.2012.03.004. [DOI] [PubMed] [Google Scholar]
- 36.Clifford HD, Hayden CM, Khoo SK, Naniche D, Mandomando IM, Zhang G, Richmond P, Le Souëf PN. Polymorphisms in key innate immune genes and their effects on measles vaccine responses and vaccine failure in children from Mozambique. Vaccine. 2012;30:6180–85. doi: 10.1016/j.vaccine.2012.07.063. [DOI] [PubMed] [Google Scholar]
- 37.[accessed 2019 Nov 8].https://bioinformatics.bethematchclinical.org/hla-resources/haplotype-frequencies/high-resolution-hla-alleles-and-haplotypes-in-the-us-population/.
- 38.Mayo O. A century of Hardy-Weinberg equilibrium. Twin Res Hum Genet. 2008;11:249–56. doi: 10.1375/twin.11.3.249. [DOI] [PubMed] [Google Scholar]
- 39.Hetherington S, Hughes AR, Mosteller M, Shortino D, Baker KL, Spreen W, Lai E, Davies K, Handley A, Dow DJ, et al. Genetic variations in HLA-B region and hypersensitivity reactions to abacavir. Lancet. 2002;359:1121–22. doi: 10.1016/S0140-6736(02)08158-8. [DOI] [PubMed] [Google Scholar]
- 40.Mallal S, Nolan D, Witt C, Masel G, Martin AM, Moore C, Sayer D, Castley A, Mamotte C, Maxwell D, et al. Association between presence of HLA-B*5701, HLA-DR7, and HLA-DQ3 and hypersensitivity to HIV-1 reverse-transcriptase inhibitor abacavir. Lancet. 2002;359:727–32. doi: 10.1016/S0140-6736(02)07873-X. [DOI] [PubMed] [Google Scholar]
- 41.Symonds W, Cutrell A, Edwards M, Steel H, Spreen B, Powell G, McGuirk S, Hetherington S. Risk factor analysis of hypersensitivity reactions to abacavir. Clin Ther. 2002;24:565–73. doi: 10.1016/S0149-2918(02)85132-3. [DOI] [PubMed] [Google Scholar]
- 42.Cutrell AG, Hernandez JE, Fleming JW, Edwards MT, Moore MA, Brothers CH, Scott TR. Updated clinical risk factor analysis of suspected hypersensitivity reactions to abacavir. Ann Pharmacother. 2004;38:2171–72. doi: 10.1345/aph.1E202. [DOI] [PubMed] [Google Scholar]
- 43.Dunstan SJ, Hue NT, Han B, Li Z, Tram TT, Sim KS, Parry CM, Chinh NT, Vinh H, Lan NPH, et al. Variation at HLA-DRB1 is associated with resistance to enteric fever. Nat Genet. 2014;46:1333–36. doi: 10.1038/ng.3143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wysocki J, Center KJ, Brzostek J, Majda-Stanislawska E, Szymanski H, Szenborn L, Czajka H, Hasiec B, Dziduch J, Jackowska T, et al. A randomized study of fever prophylaxis and the immunogenicity of routine pediatric vaccinations. Vaccine. 2017;35:1926–35. doi: 10.1016/j.vaccine.2017.02.035. [DOI] [PubMed] [Google Scholar]
- 45.Haralambieva IH, Ovsyannikova IG, Kennedy RB, Larrabee BR, Zimmermann MT, Grill DE, Schaid DJ, Poland GA. Genome-wide associations of CD46 and IFI44L genetic variants with neutralizing antibody response to measles vaccine. Hum Genet. 2017;136:421–35. doi: 10.1007/s00439-017-1768-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Schoggins JW, Wilson SJ, Panis M, Murphy MY, Jones CT, Bieniasz P, Rice CM. A diverse range of gene products are effectors of the type I interferon antiviral response. Nature. 2011;472:481–85. doi: 10.1038/nature09907. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.