
Figure 3.
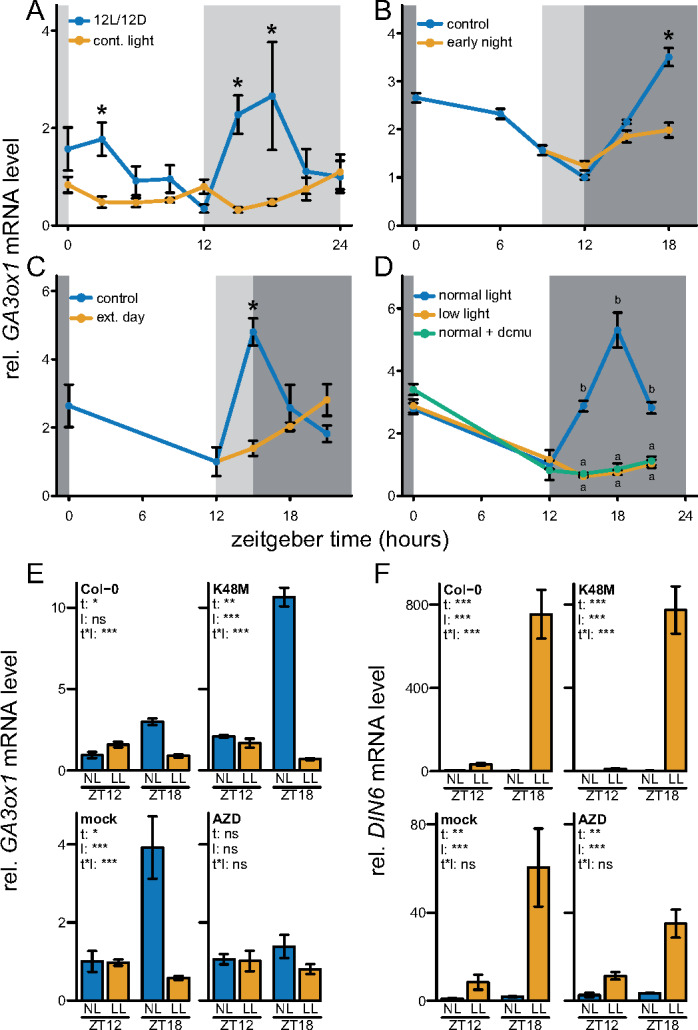
Transcriptional regulation of the nighttime peaking GA biosynthetic gene, GA3ox1. (A) The role of the circadian clock in mediating rhythmic expression. Light gray boxes represent subjective night. Sampling started second day after starting continuous light conditions. (B) Effect of a 3-h early start of the night. Dark and light gray represent night and the subjective night, respectively. (C) Effect of a 3-h extension of the day. Dark and light gray represent night and the subjective night, respectively. (D) The effect of reduced light availability or DCMU on nighttime GA3ox1 transcript abundance. DCMU was applied at the start of the preceding night. All data are from ∼10 leaf-stage, soil-grown Col-0 rosettes. Means ± SE are shown, n = 4. Asterisks (planned comparisons) and letters (Tukey HSD) represent statistically significant difference at specific time points, P < 0.05). (E, F) Manipulation of energy signaling pathways and the mRNA abundance of GA3ox1 and DIN6 at the end of the day (ZT12) and during the subsequent night (ZT18), both after control NL and LL levels. K48M: overexpresses the dominant inactive form of SnRK1.1/KIN10, where lysine residue 48 is mutated (Baena-Gonzalez et al., 2007). Blocking of TOR kinase activity by AZD8055 (AZD, 30 µM). Mock treatment (DMSO 0.01%) or AZD was applied to Col-0 at 6 a.m., 2 h prior the start of the day (ZT0, 8 a.m.). Means ± SE are shown (N = 5). Asterisks indicate the significance of effects in a two-way analysis of variance (ANOVA). *P < 0.05; **P < 0.01; ***P < 0.001. t – effect of time (ZT12, ZT18), l – effect of NL, LL, t * l, interaction of time and light.