
Figure 3:
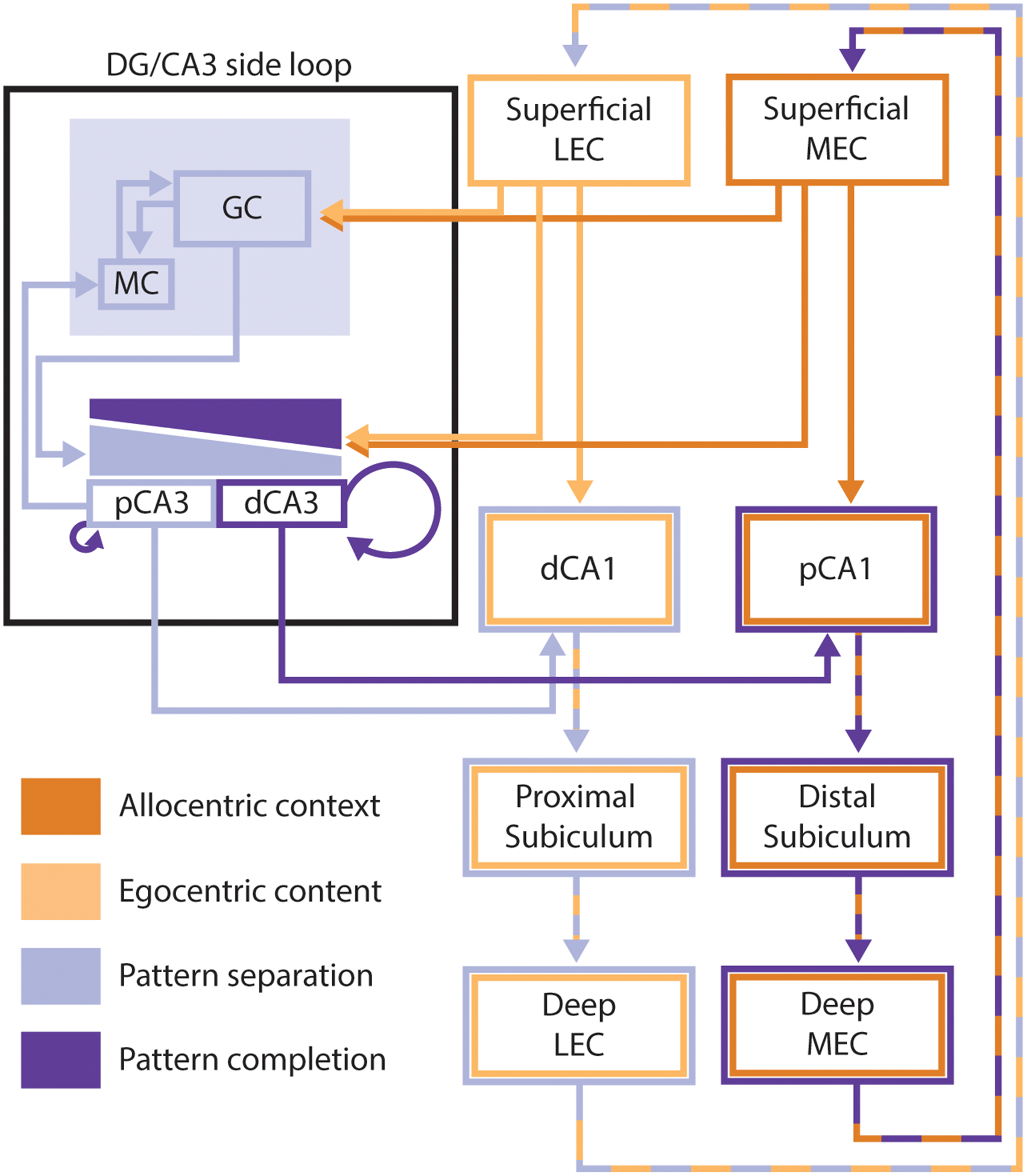
Simplified diagram of the hippocampal circuit. This diagram emphasizes the interacting parallel processing loops within the hippocampus, and how these loops interact with the complementary processes of pattern completion and pattern separation in the DG and CA3. MEC inputs reflecting allocentric contextual information (dark orange) project to distal portions of CA1, while LEC inputs reflecting the egocentric content of an experience (light orange) project to proximal portions of CA1. Both MEC and LEC project directly to the DG. The DG sends mossy fibers to CA3, with stronger inputs to the proximal than distal portion of CA3 (light purple gradient) and proximal CA3 sends backprojections back to DG mossy cells. CA3 also receives direct projections from both MEC and LEC, but this input is weaker at the proximal edge of CA3 than at more distal regions (dark purple gradient). Distal CA3 has a more extensive system of recurrent collaterals than proximal CA3 (proximal CA3 only sends collaterals to proximal CA3 while more distal regions send collaterals throughout CA3). Within the DG/CA3 side loop, the complementary processes of pattern completion and pattern separation occur. Areas and projections reflecting pattern separation (DG, proximal CA3, and communication between these areas) have been marked in light purple, while areas and projections that promote or reflect pattern completion (distal CA3, entorhinal inputs to CA3, recurrent collaterals) have been marked in dark purple. Pattern-separated outputs of proximal CA3 project preferentially to distal CA1 while pattern-completed outputs from distal CA3 project preferentially to proximal CA1. These topographical projections from CA3 to CA1 interact with the parallel input streams from the EC. The output of CA1 is then projected back to the EC (directly and via subiculum) to complete the hippocampal processing loop. Throughout the figure, proximal and distal portions of hippocampal subfields are portrayed as separate areas with entirely distinct inputs and outputs. It is important to keep in mind that, in reality, these areas exhibit gradients from proximal to distal and that the inputs, outputs, and function of these regions vary along those gradients. Further, the gradients of inputs to CA3 from entorhinal cortex and DG are highly simplified, and these gradients are not nearly as uniform as portrayed here.