
Figure 4. Intrinsic and extrinsic cues in spindle orientation and fate decisions.
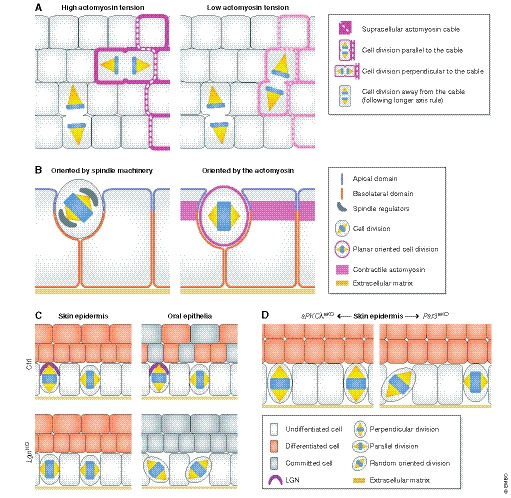
(A) In the Drosophila melanogaster germ band, spindle orientation of boundary cells is guided by the topology and anisotropy of the tensile actomyosin network, whereas inner cells not facing the boundary divide along their longer axis (Scarpa et al, 2018). (B) In the D. melanogaster follicular epithelium, spindle orientation is not coupled to tissue elongation, challenging the tissue context dependency of these mechanisms. Inhibition of mitosis in follicular cells does not affect tissue extension but impairs the optimal packing of the cells. Interestingly, in this context Pins/Mud are necessary for the apico‐basal positioning of the spindle (left) but not for its planar positioning (right) (Finegan et al, 2019). (C) In mammals, deletion of the spindle regulator LGN differentially affects mitotic spindle orientation, perhaps based on its apical vs. lateral localization in different epithelia (Byrd et al, 2016; Lough et al, 2019). In the mouse skin epidermis, LGN loss elicits mostly planar cell divisions, and in the oral tongue epithelium, it randomizes the spindle orientation. In these epithelia, LGN deficiency reduces differentiation or stratification, whereas LGN is not required for embryonic hair follicle development (Byrd et al, 2016). See text for details. (D) In the adult murine epidermis, aPKCλ deletion results in increased perpendicular divisions and epidermal differentiation (Niessen et al, 2013). Contrarily, epidermal Par3 deletion results in more planar and random divisions and increased differentiation (Ali et al, 2016).