
Figure 1. Mechanisms of innate immune signaling during alphaherpesvirus infection.
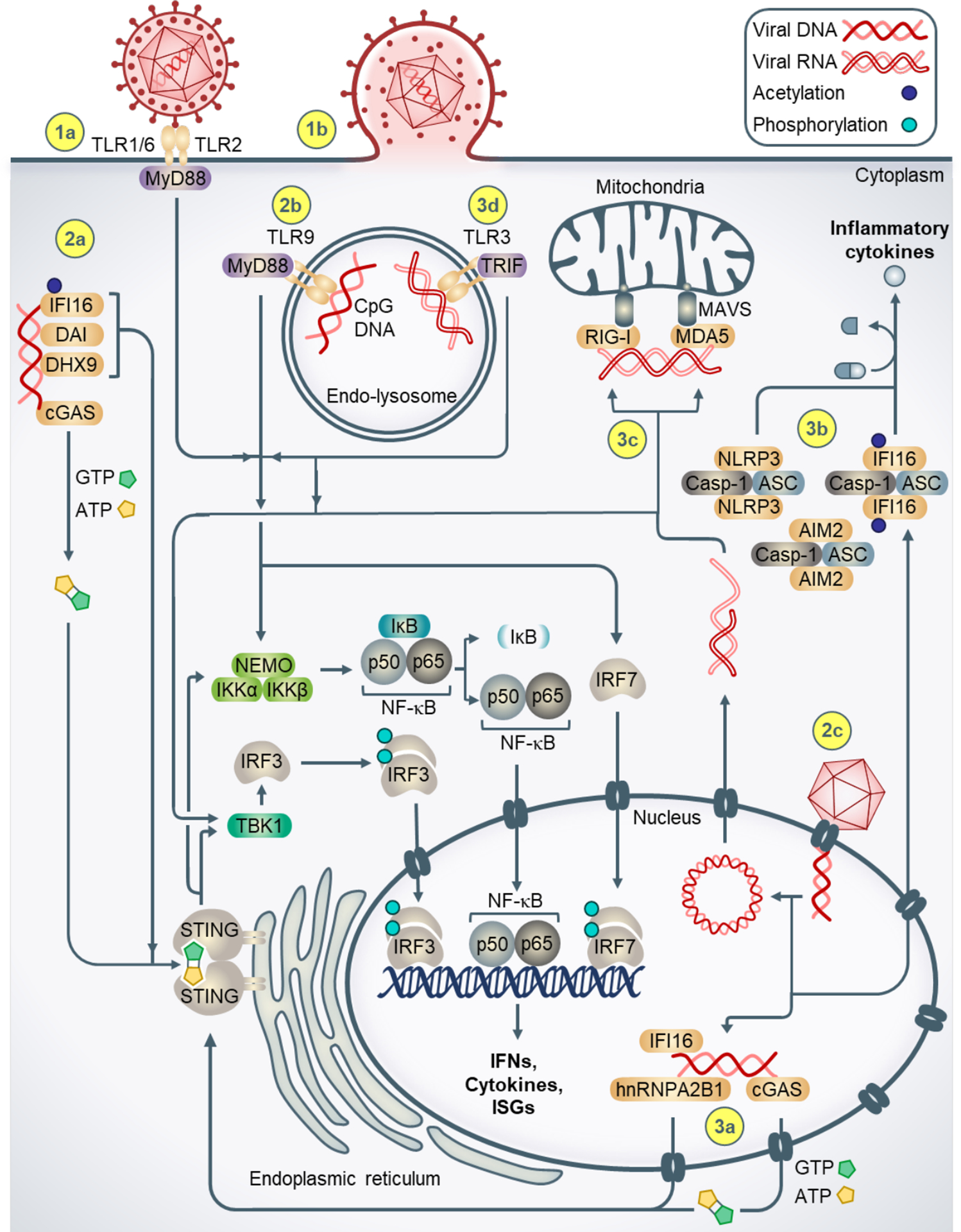
(1a) TLR2 forms heterodimers with TLR1/6, binds extracellular viral glycoproteins, and intracellularly recruits the adaptor protein MyD88. MyD88 activates the transcription factors NF-κB, IRF3, IRF7, which upregulate type I IFNs, cytokines, and ISGs. (1b) Virion fusion with the plasma membrane facilitates entry of the viral nucleocapsid and tegument proteins. (2a) Cytosolic DNA sensors bind viral DNA genomes and signal to STING at the ER. Activated cGAS generates cGAMP, which binds STING. Activated STING induces immune signaling via TBK-1 and the IKK complex, resulting in cytoplasmic to nuclear translocation of IRF3 and NF-κB and induction of type I IFNs, cytokines, and ISGs. (2b) Endosomal detection of CpG-rich viral DNA occurs via TLR9, which also recruits MyD88 to induce immune signaling. (2c) Viral nucleocapsids docking at the nuclear pore eject dsDNA genomes into the nucleus, (3a) facilitating DNA sensing by nuclear IFI16, hnRNPA2B1, and cGAS. (3b) Viral DNA-bound IFI16, NLRP3, and AIM2 inflammasomes in the cytoplasm generate pro-inflammatory cytokines. (3c) RNA intermediates generated during viral replication are sensed by RIG-I and MDA5 in the cytoplasm. (3d) Endosomal RNA is sensed by TLR3, which recruits the adaptor TRIF to activate downstream transcription factors.