
Figure 2. Mechanisms of alphaherpesvirus-mediated innate immune evasion.
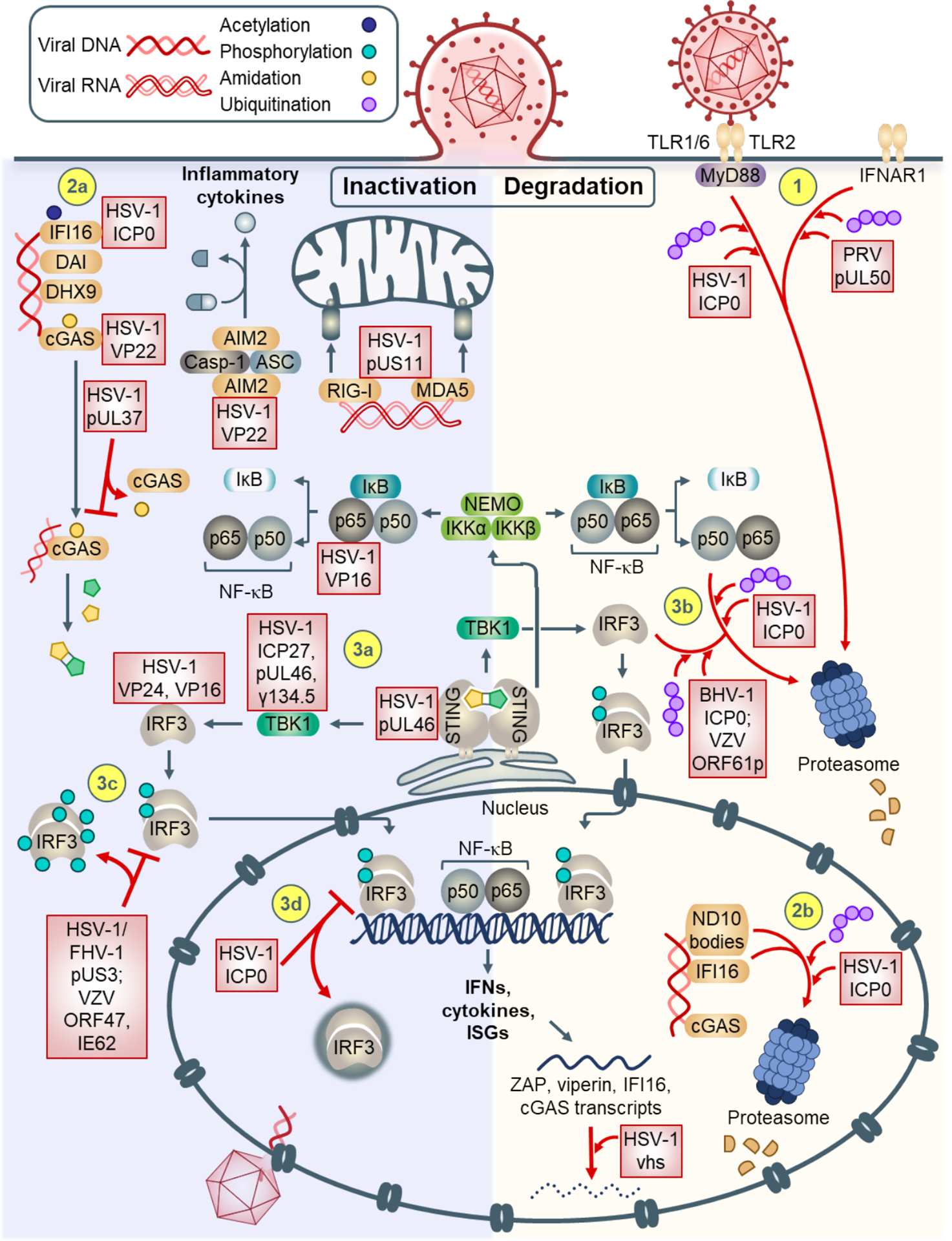
Signal transduction can be inhibited via inactivation (left), or degradation (right) of immune response components. (1) At the plasma membrane, IFNAR1 and TLR2-recruited MyD88 can be targeted for proteasomal degradation. (2a) Cytosolic DNA sensors IFI16 and cGAS are inhibited by direct binding. (2b) In the nucleus, ND10 body components and IFI16 are targeted for proteasomal degradation via HSV-1 ICP0. Upon activation of the STING-TBK-1-IRF3 pathway, (3a) STING, TBK-1, IRF3, and NF-κB p65 are inhibited by alphaherpesvirus proteins via binding. (3b) Additionally, IRF3 and NF-κB p50 can be targeted for degradation. (3c) IRF3 can also be atypically phosphorylated, preventing proper dimerization and nuclear translocation. (3d) HSV-1 ICP0 can also sequester IRF3 into nuclear puncta, preventing its transcriptional activity. (4) Degradation can occur at the RNA level (e.g., ISG transcripts ZAP, viperin, IFI16, cGAS). (5) Sensors AIM2, RIG-I, and MDA5 can be inhibited by binding of HSV-1 proteins.