
Figure 1.
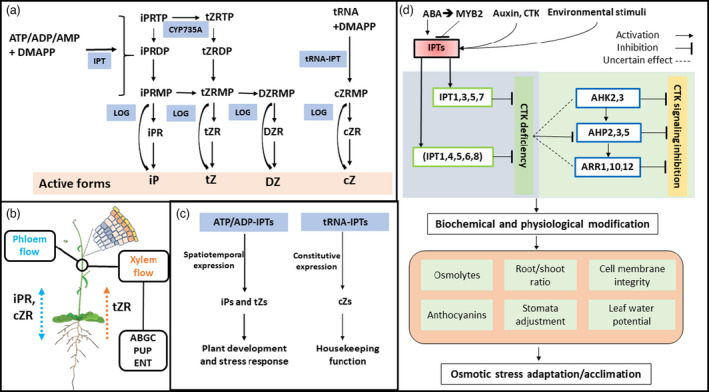
An overview of CTK biosynthesis (a); CTK translocation (b); general characteristics of two known CTKs biosynthesis pathways in angiosperms (c) and, the mode of action of IPTs in drought tolerance in Arabidopsis (d). (a) CTK types derived from two known pathways, adenylate (ATP/ADP‐IPTs and AMP‐IPTs) and tRNA types (adapted from Hirose et al. (2008), further details in Sakakibara (2006)). ADP: adenosine 5’‐diphosphate; ATP: adenosine 5’‐triphosphate; tRNA: Transfer ribonucleic acid; DMAPP: dimethylallyl diphosphate; IPT: isopentenyl transferase; CYP735A: cytochrome P450 monooxygenase, family 735, subfamily A (cytokinin trans‐hydroxylase); LOG: cytokinin phosphoribohydrolase ‘Lonely guy’; cZ: cis‐Zeatin, cZR: cZ riboside; cZRMP: cZ riboside 5’‐monophosphate, DZ: Dihydrozeatin, DZR: DZ riboside; DZRMP: DZ riboside 5’‐monophosphate; iP: isopentenyladenine; iPR: iP riboside; iPRMP: isopentenyladenosine‐5’–monophosphate; iPRDP: isopentenyladenosine‐5'‐diphosphate; iPRTP: isopentenyladenosine‐5'‐triphosphate; tZ: trans‐Zeatin, tZR: tZ riboside, tZRDP: tZ riboside 5’‐diphosphate; tZRMP: tZ riboside 5’‐monophosphate; tZRTP: tZ riboside 5’‐triphosphate. (b) Translocation of CTKs in plant. As multifunctional and mobile signalling molecules, active CTKs and their derivatives contribute to various developmental processes depending on CTK transport proteins across vascular tissues. Briefly, three kinds of CTK transporters have been systematically characterized: equilibrate nucleoside transporters (ENT), purine permeases (PUP), and G subfamily ATP‐binding cassette (ABCG) transporters (Durán‐Medina et al., 2017). tZR is the primary form of xylem transported CTKs, and iPR and cZR are major forms of phloem CTKs (Osugi and Sakakibara, 2015). Arabidopsis plant vector is adapted, with permission, from Figshare (Bouché, 2018). (c) General mode of action of two IPT GFMs (ATP/ADP‐IPTs and tRNA‐IPTs) in angiosperms (Köllmer et al 2014; Miyawaki et al 2006; Wang et al., 2020a). The investigation in Arabidopsis mutants showed that ATP/ADP‐IPT GFMs control biosynthesis of iP‐ and tZ‐type CTKs while tRNA‐type IPT genes regulate cZ‐type CTKs (Köllmer et al., 2014; Miyawaki et al., 2006). In the basal angiosperm Amborella trichopoda (Amborellaceae), and in Fragaria vesca (wild strawberry), eudicot woodland strawberry, transcriptomic analysis indicated that tRNA‐IPTs are constitutively expressed throughout the plant, whereas the expression of ATP/ADP‐IPTs is tissue‐specific and rapidly down‐regulated by abiotic stresses (Wang et al., 2020a). Generally, tRNA‐IPTs and the associated cZ‐type CTKs play a housekeeping role, whereas ATP/ADP‐IPTs and associated iP/tZ‐type CTKs play regulatory roles in organ development and stress responses in angiosperms (Köllmer et al., 2014; Miyawaki et al., 2004, 2006; Wang et al., 2020a). (d) The negative regulatory role of IPT‐repressed CTKs in osmotic stress tolerance in Arabidopsis. Arrowheads represent activation, and perpendicular bars indicate inhibition. Studies of drought stress tolerance using CTK‐deficient plants, such as the quadruple ATP/ADP‐ipt1,3,5,7 loss‐of‐function mutants, CTK signalling mutants [AHP mutants (Arabidopsis Histidine Phosphotransfer Proteins 2,3,5) and ARR mutants (type B Arabidopsis Response Regulators 1,10,12)], and down‐regulated expression of five ATP/ADP‐IPT genes (IPT1, IPT4, IPT5, IPT6, IPT8) found in the AtMYB2 (ABA‐dependent signalling pathway) overexpressor, have indicated that CTK‐depleted mutants have improved drought acclimation/adaptation (Guo and Gan, 2011; Nguyen et al., 2016; Nishiyama et al., 2011, 2013; Werner et al., 2010). Briefly, reducing endogenous CTK lowers the output of the CTK signalling cascades (i.e. quadruple ipt1,3,5,7 down‐regulates expression of AHP2, AHP3, AHP5) leading to drought acclimation/adaptation. CTK: cytokinin; AHK: Arabidopsis histidine kinase; AHP: Arabidopsis histidine phosphotransfer; ARR: Arabidopsis response regulator; ROS: reactive oxygen species.