
Abb. 3.
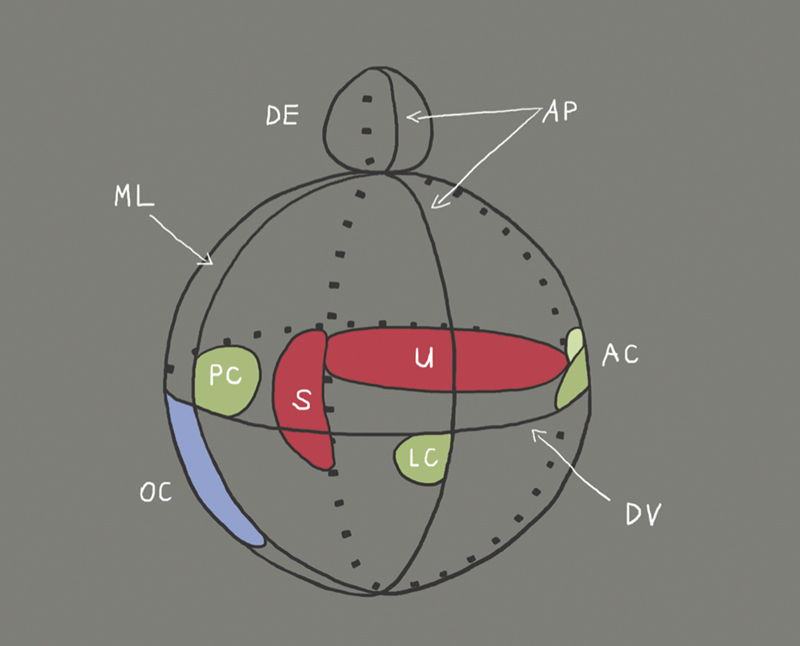
Schematische Darstellung der Kompartimente des sich entwickelnden Innenohres und Lokalisation der verschiedenen Organe (Corti-Organ: OC, Sacculus: S, Utriculus: U, Ductus endolymphaticus: ED, Cristae der Bogengänge: AC, PC und LC) sowie der Achsen (AP: antero-posterior; DV: dorso-ventral; ML: medio-lateral). Die Ausrichtung in der antero-posterioren Achse erfolgt vor der Ausrichtung nach dorso-ventral 26 . Die dorsoventrale Achse ist bis zur Ausbildung der Otozyste noch nicht festgelegt 27 . Die Achsenspezifikation beginnt bereits mit der Ausbildung der otischen Plakode und hängt von Faktoren ab ( Abb. 2 ), die von den Rhombomeren 5 und 6 des Rautenhirns exprimiert werden. Sobald ein Rhombomer entlang der dorso-ventralen Achse in ovo rotiert ist, verschiebt die Expression der ventralen Gene Lfng, NeuroD1 und Six1 (siehe Tabelle BOR-Syndrom) in die dorsalen Regionen der Otozyste, wohingegen die Expression dorsaler Gene wie Gbx2 gehemmt ist. Dies bedeutet, dass durch Rotation der Rhombomere ventrale Bezirke des Rautenhirns ventrale Bereiche der Otozyste in dorsales otisches Gewebe transformieren können 26 . Die Ausbildung der Organe des Innenohres nach dem Stadium der Otozyste ist von der Expression von Gata3 (Siehe Tabelle Bakarat-HDR-Syndrom), Eya1 (siehe Tabelle BOR-Syndrom) und FGF3 / 8 (siehe Tabelle Kallmann-Syndrom) abhängig, wie Untersuchungen an Gata3-, Eya1- und FGF3/8-defizienten Mäusen gezeigt haben 28 29 30 . SHH (siehe Tabelle Inkomplette Partition und Carpenter-Syndrom) sowie Pax2 sind cochleäre Gene, da Mutationen in diesen Genen, lediglich die Ausbildung eines kurzen, geraden Ductus cochlearis erlauben. Als vestibuläre Gene werden hingegen Gbx2, Hmx2, Hmx3 sowie WNT bezeichnet, da ein Ausfall einer dieser Gene zu morphologischen Defekten in Sacculus, Utriculus oder den Bogengängen führt. (Zeichnung modifiziert nach Brigande et al., 2000 16 .)