
FIGURE 3.
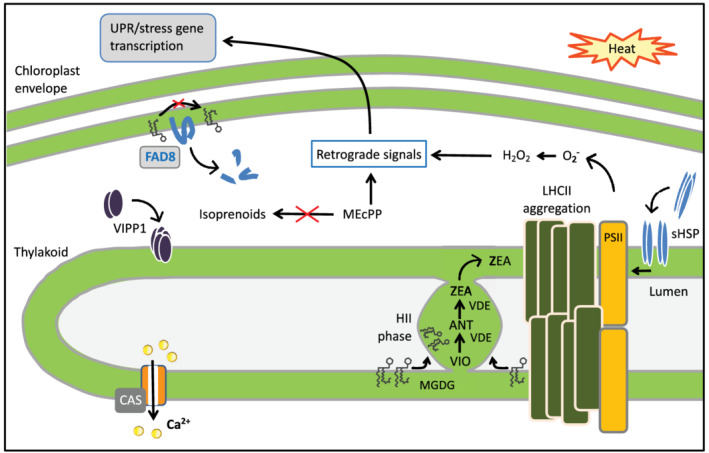
Sensing and signalling of heat stress at the chloroplast. Exposure to moderate heat has various direct consequences for chloroplast proteins and membranes, which trigger rescue pathways. In the thylakoidal membrane, LHCII proteins aggregate, which leads to an excess of its major lipid constituent, the non‐bilayer prone glycerolipid MGDG. As a consequence, a non‐bilayer structure (the HII phase) emerges, which consists of MGDG organized in hexagonally stacked tubules. The xanthophyll cycle enzyme, violaxanthin de‐epoxidase (VDE), recruits specifically to the HII phase in the thylakoid lumen, catalysing the synthesis of zeaxanthin (ZEA) from its precursors, violaxanthin (VIO) and antheraxanthin (ANT). The HII phase remains attached, which allows for free diffusion of the photoprotective xanthophylls to the thylakoid. VIPP1 and sHSP recruit to the thylakoid membrane under heat stress, as they recognize membrane packing defects. They protect thylakoid membrane and PSII integrity. Heat stress is signalled in the chloroplast by a rapid Ca2+ increase in the stroma which depends on the activity of the calcium sensor CAS. Furthermore, heat induces breakdown of the envelope desaturase FAD8, responsible for synthesis of polyunsaturated fatty acids. This causes an adaptive decrease in membrane desaturation. The isoprenoid biosynthesis intermediate MEcPP accumulates due to a high light/temperature‐induced bottleneck in the pathway. MEcPP, together with H2O2 resulting from excess excitation energy, and other stress‐induced molecules, serve as retrograde signals to regulate heat stress response genes in the nucleus