
FIGURE 1.
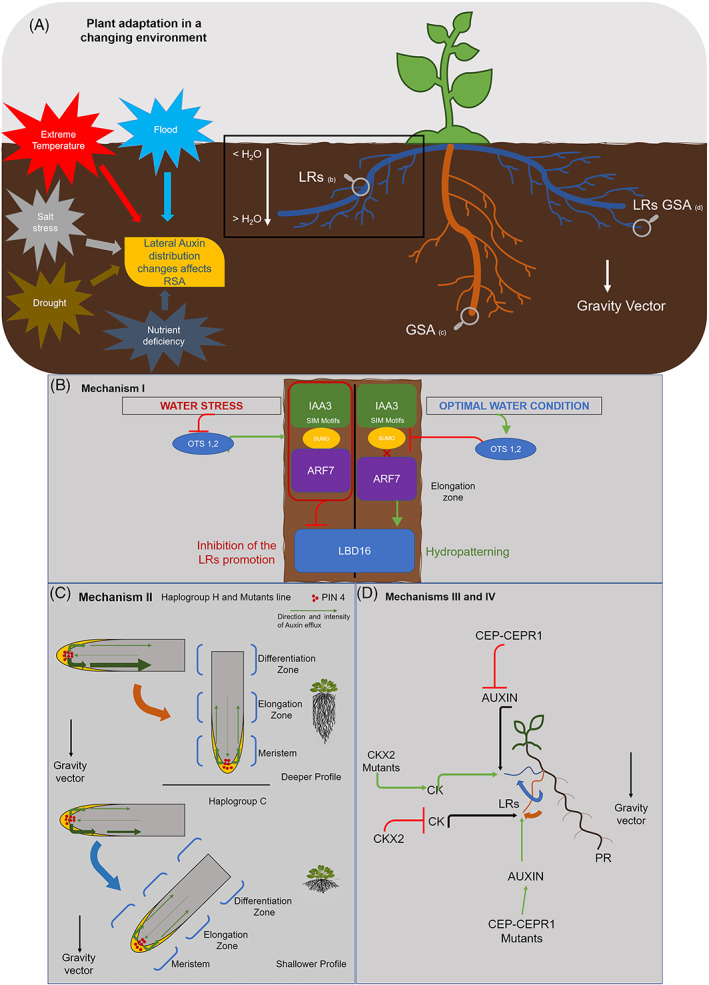
Molecular mechanisms driving root system architecture (RSA). (A) RSA is the phenotypic result of the interaction of genetic and environment controls (G × E)—Abiotic stresses (e.g. flooding; extreme temperatures; salt stress, etc.), often exacerbated by climate change, affect lateral auxin distribution via multiple molecular mechanisms, highlighted by the magnifier glasses, and elucidated in the following panels. (B) Root branching (lateral root [LRs] promotion) is shaped by mechanism I—Hydropatterning affects root branching in the elongation zone of the primary root (PR). Environmental condition, i.e. water stress and optimal water conditions affect SUMO proteases (OTS1,2). On the dry side of the root (on the left of Panel B), water stress inhibits SUMO proteases, in turn allow the formation of the repressor complex (red circle) (IAA3‐SUMO‐ARF7), thereby blocking (red arrow) auxin‐responsive gene expression associated with LR initiation. On the right of Panel B (optimal water conditions), moisture inhibits (red arrow) the formation of the repressor complex (IAA3‐SUMO‐ARF7) affecting SUMO. Thus, ARF7 induced asymmetric expression of LBD16 in LRs. Light green arrow: activation; red arrow: inhibition; red cross: missing contact between ARF7 and SUMO. (C) Gravitropic set‐point angle (GSA) is shaped by mechanism II—Green arrows show the direction and intensity of auxin efflux via PIN4 (red points). The exocyst factor, EXO70A3, modulate the asymmetric auxin gradient between the lower and upper side of the root tip via PIN4 proteins. Gravity stimulus in columella cells sets the direction of the root bending according to the allelic variation of EXO70A3 determining the threshold of auxin flux. Thus, two models of extreme wild‐type phenotypes resulting in a deeper RSA profile (upper side) and in a shallower RSA profile (lower side) are proposed. Shallower RSA (in blue) is shared by haplogroup C and guarantees exploration of the soil more shallower layers and access to superficial water by herbaceous crops. However, some selected varieties, like haplogroup H, root deeply (in orange). (D) GSA of LRs is shaped by mechanisms III and IV—CEP‐CEPR1 and cytokinin (CK) signalling are proposed as molecular mechanism shaping LRs GSA during the III stage. CEP‐CEPR1 signalling inhibits both rootward auxin transport and shoot auxin levels, in turn affecting bending of GSA of the LRs and inducing a shallower root profile (blue arrow). Auxin flux that affects GSA of the LRs is balanced by CK that act as anti‐gravitropic signal at opposing organ flanks. CK oxidase 2 (CKX2) affects CK pathway, its natural allelic variation shapes different degree of LRs bending. CKX2 mutants and wild type reflect two opposite phenotypes respectively showing a shallower and deeper LRs profile. Light green arrow: activation; red arrow: inhibition; black arrow: a consequence of a previous activation or inhibition