
Figure 2. The origin and diversification of a new sex-specific trait.
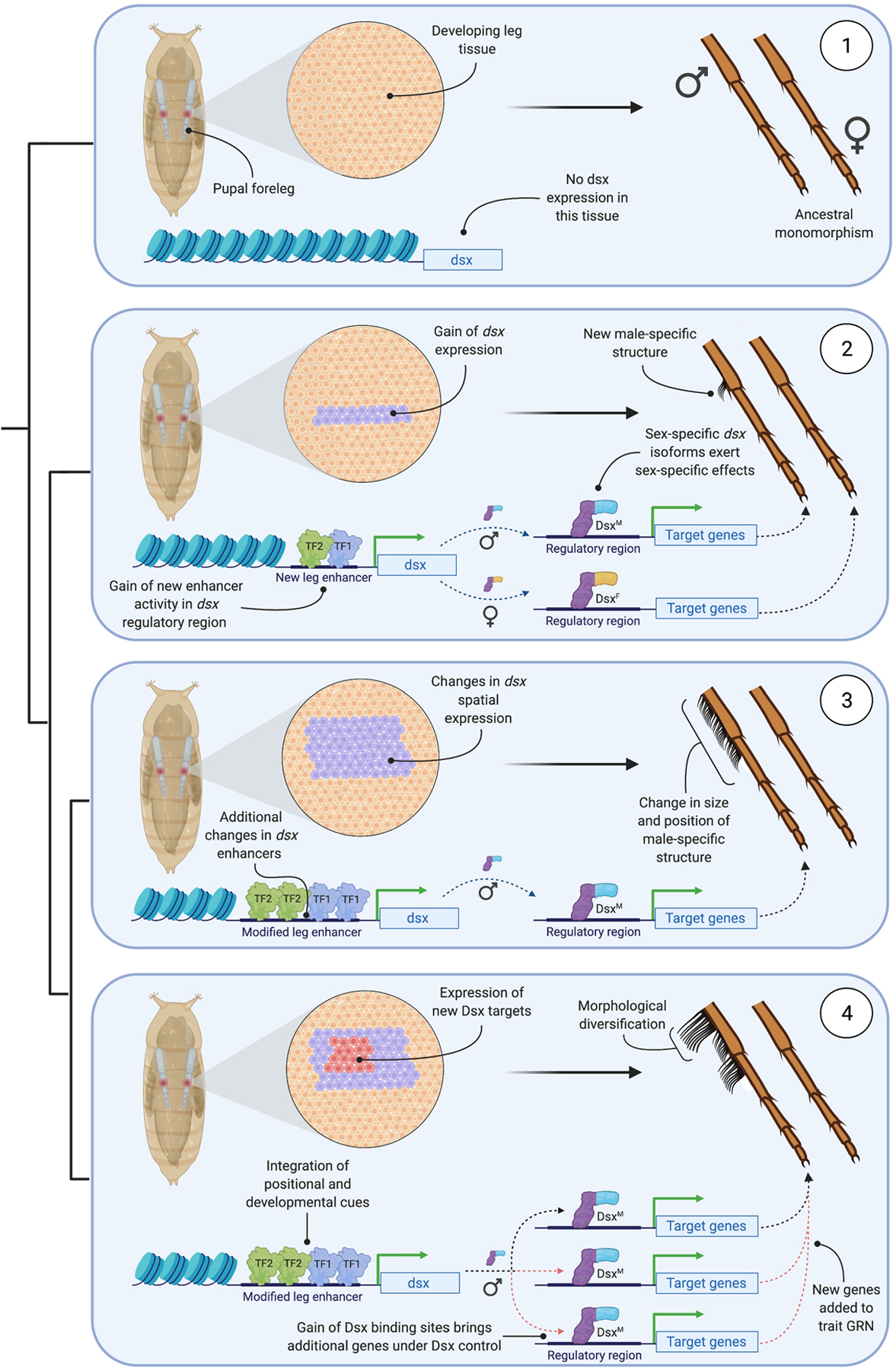
This schematic describes a four-part model for the origin and subsequent morphological diversification of a sex-specific structure, in this case a modified row of bristles (a ‘sex comb’) on the male Drosophila foreleg. Species 1 displays the ancestral state of monomorphism. Here, developing leg cells do not express the transcription factor doublesex (dsx) and therefore lack the capacity for sex-specific differentiation. In species 2, changes in the sequence of the regulatory region controlling dsx expression enable the binding of position- and stage-determining transcription factors (TF). These TFs activate dsx expression in a subset of leg cells during a particular developmental window. dsx is alternatively spliced to give rise to male- and female-specific isoforms (DsxM and DsxF), which bind to the regulatory regions of target genes via a shared DNA-binding domain and impart sex-specific effects on target gene expression through sex-specific C-termini. The localized, sex-specific regulation of gene expression that results enables the development of a novel structure only in males. In species 3, additional changes in the dsx enhancers generate changes in the binding of its upstream regulators. This leads to changes in the spatiotemporal pattern of dsx expression among developing leg cells, which in turn produces changes in the size and position of the male-specific structure. In species 4, Dsx has acquired new downstream target genes due to the gain of Dsx-binding sites in the regulatory regions of the new targets. Incorporation of the new targets into the gene regulatory network (‘GRN’) that controls the development of the male-specific structure leads to further morphological diversification. Figure created using BioRender.