
FIGURE 1.
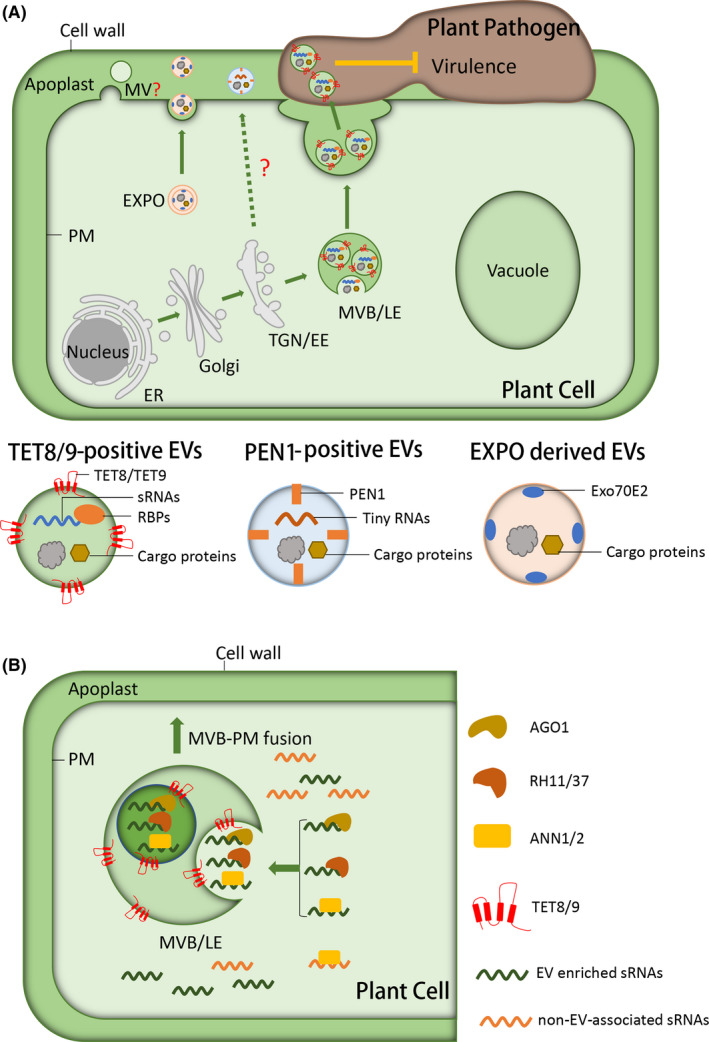
Plant EVs communicate with interacting microbes. A, A heterogeneous population of EVs exists in plants. After fusion with PM, MVBs release TET8/9‐positive EVs into the extracellular space. TET8/9‐positive EVs contain defense‐related molecules such as host‐derived sRNAs, defense proteins, and RBPs that function in sRNA sorting and stabilization in EVs. These host‐derived defense molecules can be internalized by plant pathogens to suppress pathogen virulence. Another class of EVs is PEN1‐positive EVs, which contains tiny RNAs. But the biogenesis pathway of PEN1‐positive EVs and whether they contribute to plant immune responses or cross‐kingdom regulation is not yet clear. EXPO produces another class of single‐membrane‐bound EVs, which may aide in the re‐localization of defense‐related molecules during pathogen invasion. MVs may also be secreted by plant cells through outward budding directly from the PM. The three major classes of plant EVs are presented in the bottom panel individually. The question mark indicates a prediction that has not yet been validated experimentally. B, RNA‐binding proteins contribute to sRNA loading into plant extracellular vesicles. Arabidopsis Argonaute 1 (AGO1), RNA helicase (RH) 11, and RH37 selectively bind to EV‐enriched sRNAs but not to non‐EV‐associated sRNAs, annexin (ANN) 1 and ANN2 bind to sRNAs non‐specifically. These RNA‐binding proteins with EV‐enriched sRNAs are targeted for capture into a bud invaginating into the MVB. ER, endoplasmic reticulum; EV, extracellular vesicle; EXPO, exocyst‐positive organelle; LE, late endosome; MV, microvesicle; MVB, multivesicular body; PEN1, Penetration 1; PM, plasma membrane; RBPs, RNA‐binding proteins; TET, Tetraspanin; TGN/EE, trans‐Golgi network/early endosome