

Abstract
In recent years, the H9N2 influenza virus has been circulating widely in poultry farms causing extensive damage. The hemagglutinin (HA) genes of the two virus isolates of H9N2 subtype in specific pathogen-free chickens were studied to determine the shedding rate in the host’s oropharyngeal and cloacal routes and their genetic relationship. The sequence analysis and phylogenetic study of the samples were performed by comparing each isolate with other H9N2 isolates in the gene bank. In the present study, the chickens were inoculated with low pathogenic avian influenza virus (LPAIV) (A/Chicken/Iran/ZMT-101/1998 [H9N2]) through the intranasal route. Oropharyngeal and cloacal swabs were collected from the chickens within 1-10 days after inoculation. The rate of viral shedding was measured within the previous 10 days by the real-time reverse transcriptase polymerase chain reaction molecular technique. No clinical symptoms were observed during the experiment in the chickens. The results obtained from this technique showed that the main route of shedding for LPAIV was oropharyngeal areas (p<0.05). Both isolates had a similar proteolytic R-S-S-R sequence at the cleavage site of the HA gene and contained glutamine (Q) amino acid at position 226 of the HA receptor-binding site, indicating that these isolates were nonpathogenic. Phylogenetic analysis demonstrated that both isolates belonged to the Eurasian clade. The comparison of these isolates with other isolates in the gene bank showed that they had the greatest similarity with the isolates in clade 1 and the least homology with the isolates in clade 4.
Keywords: Influenza virus, H9N2, Real-time RT-PCR, Phylogenetic analysis
Introduction
Avian influenza is a highly contagious and acute respiratory system disease that causes serious economic damages to the poultry industry ( Karimi et al., 2015 ). Avian influenza virus (AIV) is classified under the Orthomyxoviridae family, and its genome is a single-stranded ribonucleic acid (RNA), segmented, circular, and polymorphic, with negative-sense ( Pazani et al., 2008 ). The influenza virus’s genome encodes 10 types of protein. Eight types of these proteins, including hemagglutinin (HA), M1, M2, NP, PA, NA, PB1, and PB2, are structural proteins of the virus, and two types, namely NS1 and NS2, are nonstructural. Based on the antigenic characteristics of surface glycoproteins of HA (i.e., H1 to H17) and neuraminidase (i.e., N1 to N9), type A influenza viruses are divided into different subtypes ( Fouchier et al., 2005 ). The AIVs are divided into two general categories of highly and nonhighly avian influenza pathogenic viruses ( Vasfi Marandi et al., 2002 ; Pazani et al., 2008 ). The surface glycoprotein of the influenza viruses plays an important and fundamental role in the infectious processes of the virus. It is also responsible for the virus binding to the sialic acid receptor cells on the host cell surface and facilitates the virus entry into the host cell by endosomal membrane fusion ( Zhu et al., 2013 ; Costa-Hurtado et al., 2014 ). Low pathogenic avian influenza viruses (LPAIVs) cause mild clinical signs and some subclinical infections ( Lee et al., 2007 ). The LPAIVs are typically observed in both the feces and respiratory secretions of chickens ( Lee et al., 2007 ). Few reports have described that certain routes for the shedding of LPAIV are oropharyngeal; therefore, LPAIVs are shed mainly in the respiratory secretions ( Costa-Hurtado et al., 2014 ). The sequences of amino acids at the cleavage site of the HA gene determine the viral division characteristics ( Hosseini et al., 2017 ). The presence of leucine amino acid at position 226 of HA protein provided the virus with the possibility of detecting SAα2,6 receptors in human cells ( Matrosovich et al., 2000 ). Therefore, in case of virus circulation in Iranian poultry farms, the outbreak of point mutation in these viruses is possible ( Toroghi and Momayez, 2006 ; Karimi et al., 2015 ). In 1998, the H9N2 subtype caused severe contagion, outbreak of the disease, and extreme economic damages in Iran, which were detected and reported for the first time by Vasfi Marandi et al. (2002) as A/Chicken/Iran/259/1998. Consequently, through the spread of severe and contagious diseases, it has brought about severe damages to poultry farming ( Vasfi Marandi et al., 2002 ). Given the influenza involvement, there is a span between the onset of the disease and final diagnosis; during this time, the virus might get out of control and infect large species of birds. Moreover, considering the spread of the disease in the neighboring countries, particularly Turkey, Iraq, United Arab Emirates, and Saudi Arabia, it is essential to constantly control the virus characteristics leading to the disease ( Monne et al., 2008 ; Wan et al., 2008 ). Although the influenza symptoms are similar to those of other respiratory infections, such as Newcastle disease and bronchitis, the diagnosis of the disease based on autopsy and clinical symptoms is difficult; therefore, there is a need for the development of methods capable of quick diagnosis and detection ( Liolios et al., 2001 ). A quick diagnostic method is real-time reverse transcriptase polymerase chain reaction (rRT-PCR) ( Spackman et al., 2002 ; Bashashati et al., 2013 ) . In this study, we evaluated the shedding of the Iranian strains of influenza virus (i.e., H9N2) in the oropharyngeal and cloacal routes of the specific pathogen-free (SPF) chicks using the rRT-PCR molecular technique. Furthermore, the genetic changes of the cleavage site in the aforementioned virus were analyzed with other influenza viruses, and a phylogenetic study was carried out.
Material and Methods
Virus strains
In the present study, A/Chicken/Iran/ZMT-101/1998 (H9N2), which is an isolate obtained from the infected birds in Iran, was provided by Razi Vaccine and Serum Research Institute South of Iran Branch (Shiraz).
Sample duplication and preparation
The H9N2 subtype virus was inoculated to 15 SPF 28-day-old chicks. The approval for this study was achieved from the Animal Research Ethics Committee of Islamic Azad University, Shiraz, Iran (No. 1395.145826; 2016/06/05). In this age group, Maternal Derived Antibodies titer is at its lowest, and the host is at the highest risk of developing a viral infection. Each chicken received 50 μl of the virus at a rate of 106EID50/100 µl through the intranasal route. All the chicks were daily monitored to observe symptoms. For molecular investigation and tests, cloacal and oropharyngeal swabs were taken within the first to tenth day after inoculation. All the swab samples were kept in a phosphate-buffered saline (PBS) sterile solution, and they were passed through a 0.22-μm syringe filter and kept at -70°C until RNA extraction to ensure noncontamination.
RNA extraction
Cloacal and oropharyngeal swabs were collected from all the chickens within the first to tenth day after inoculation for the evaluation of viral shedding. The samples were stored in a PBS-Bovine serum albumin liquid with an antibiotic content of specific concentrations, including penicillin G (2000 u ml-1) and gentamicin (200 μg ml-1) in -70°C up to the end of the test process. To extract virus RNA from the collected samples, the NucleoSpin® RNA kit (Macherey-Nagel GmbH & Co., Germany) was used according to the proposed instruction. This stage of the experiment was carried out at the Istituto Zooprofilattico Sperimentale delle Venezie in Italy.
Design of Probes and Primers
For the detection of H9N2 in a molecular process, the nucleotide sequences of the matrix (M) gene were considered for the influenza virus. Therefore, specific primers and probes were designed for the detection of the M gene of H9N2. The used oligonucleotide sequences of the primers and specific probes are presented as follows:
M+25 5´- AGA TGA GTC TTC TAA CCG AGG TCG -3´
M-124 5´- TGC AAA AAC ATC TTC AAG TCT CTG - 3´
M+64 [FAM] 5´- TCA GGC CCC CTC AAA GCC GA - 3´ [TAMRA]
All the probes are labeled at the 5´ end with the 6-carboxyfluorescein (FAM) reporter dye and 3´ end with the 6-carboxytetramethylrhodamine (TAMRA) quencher dye ( Spackman et al., 2002 ). Whatever was used for the detection of AIV had been discussed in previous studies ( Spackman et al., 2002 ; Monne et al. (2008) ).
Real-time RT-PCR
The reagents contained in a Quanti Tec multiplex RT-PCR kit (Qiagen, Hilden, Germany) were used for rRT-PCR. The primers targeting the M gene of AIV were applied to PCR at the optimized concentration of 300 nM. Specific fluorescently labeled probes were used at a final concentration of 100 nM for AIV. The rRT-PCR took place in the final volume of 25 µl using a Rotor-Gene 6000 apparatus (QIAGEN's real-time PCR cycler, the Rotor-Gene Q, QIAGEN Company, Netherlands). Each PCR tube contained a single primer/probe set for AIV. The identical thermal profile was adapted in order to detect viruses. The protocols used for M gene primers/probes set were 20 min at 50 ºC and 15 min at 95 ºC, followed by 40 cycles at 94 ºC for 45 sec and 60 ºC for 45 sec.
cDNA Synthesis
To synthesize complementary deoxyribonucleic acid, the RNA extracted from the SuperScript ш First-Strand Synthesis System (Thermo Fisher Scientific, USA) was used. Accordingly, the H9 gene primer of the influenza virus (RF671) with 5-AGCAAAAGCAGG-3́ sequence was also utilized according to the protocol proposed by the manufacturer. This reaction’s incubation was carried out at 25 °C for 5 min and then at 50 °C for 60 min. Afterward, a PCR reaction was performed with a 25-μl total volume. Subsequently, to amplify the desired product, PCR reaction’s temperature was adjusted as Ampli Taq activation temperature of 95 °C for 5 min in 90 cycles, including denaturation phases with a temperature of 94 °C for 30 sec, annealing phase with a temperature of 50 °C for 30 sec, and extension phase with a temperature of 72 °C for 2 min. Finally, the final extension was performed at a temperature of 72 °C for 3 min.
Sequencing and Phylogenetic Analysis
In this study, the determination of the nucleotide sequence of viral specimens was conducted using specific primers designed by the Istituto Zooprofilattico Seprimentale delle Venezie through Sanger sequencing. All the data related to the nucleotide sequence were edited by DNASTAR software (Burland 2000). The piece that could be translated into the protein of the aforementioned genes was aligned with other genes in the gene bank using online Maff software (version 7.0). Finally, the phylogenetic tree was drawn using the MEGA 7 software through maximum likelihood phylogenetic trees based on a study by Tamura et al. (2013).
Results
Clinical Symptoms
Throughout the experiment, no signs and symptoms of the disease were detected in the hosts.
Infection and viral shedding
The rate of viral shedding in the cloacal and oropharyngeal swabs was evaluated by the rRT-PCR technique. In all the oropharyngeal and cloacal swab samples, the M gene of the matrix was evaluated for H9N2 virus infection. By the comparison of the viral shedding via the rRT-PCR technique at different times after inoculation, the shedding pattern in the oropharyngeal swab showed that the rate of virus shedding in the oropharyngeal swab was at its highest within the 1st to 4th day after inoculation and gradually declined; accordingly, after the 6th day, shedding was not reported in the oropharyngeal swab. However, the rate of virus shedding in the cloacal swab was reported to be distinctly low in the early days. The evaluation of the rate of influenza virus shedding in the cloacal swab using the rRT-PCR technique indicated that the highest virus shedding was within the 2nd to 6th day after the insemination, and no shedding was observed in the cloacal swab on the 7th day (P<0.05; Figure 1).
Figure 1.
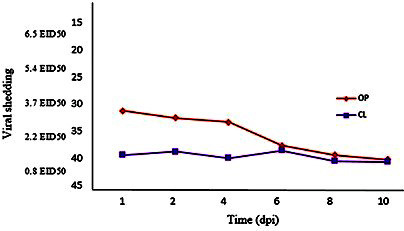
Each data point and equivalent EID50/µl representing titers of low pathogenic avian influenza virus detected in oropharyngeal and cloacal swabs at different time points after inoculation showing viral shedding of single infected birds.
According to the Applied Biosystems recommendations, detection efficiency between 98-100% indicates the optimal value. Accordingly, the results of the present study showed that efficiency was 99%; therefore, the assay was optimal. The standard curves generated from the amplification plots demonstrated a linear correlation between Ct values and virus dilutions. The presently developed assay was also shown by the slope value as -3.19, and the assay R2 value was reported as 0.997 (Figure 2).
Figure 2.
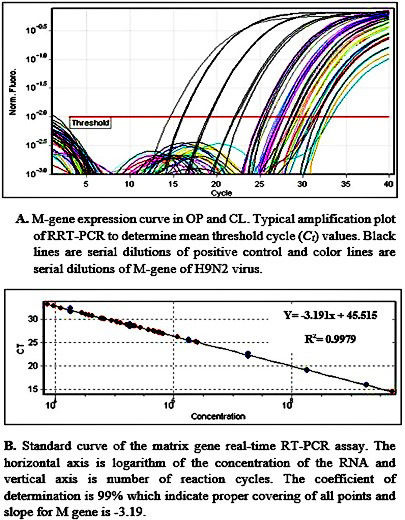
Real-time polymerase chain reaction amplification of M gene of H9N2 virus; A) M gene expression curve in oropharyngeal and cloacal swabs; B) standard curve derived from the amplification of the allantoic fluid containing avian influenza virus.
Analysis of the nucleotide sequence of HA:
Gene Bank accession number
The desired samples’ PCR product was sequenced by specific primers designed by Sanger sequencing. These sequences were recorded in the gene bank with MF114292 and MF114293 accession numbers for D1 (i.e., the samples taken on the 1st day following inoculation) and D7 (i.e., the samples taken on the 10th day after inoculation) samples, respectively.
Sequencing analysis
The sequenced part of the HA gene in the two studied viral samples contained 1618 nucleotides encoding amino acids. This protein region consists of the entire receptor binding site and cleavage site. The nucleotide sequences of these two isolates were compared with other pre-sequenced H9N2 strains and stored in the gene bank. The position of each amino acid was numbered according to the HA gene sequence under human H3 strain A/Aichi/2/68 isolates. The comparison of the nucleotides sequencing of these isolates with A/Turkey/Wisconsin/66 prototype showed no nucleotide elimination or increase in these parts. It should be pointed out that the amino acids in the receptor-binding site and cleavage site of the HA gene had distinct differences. The current study examined the amino acid positions of 183, 190, 226, and 227 of the receptor-binding site and 326, 327, 328, and 329 of the cleavage site. The sequence of amino acids in the HA gene cleavage site has an R-S-S-R motif. Both D1 and D7 isolates have similar amino acids at 326 R (arginine), 327 S (serine), 328 S (serine), and 329 R (arginine) for the cleavage site and the same amino acids at 183 H (histidine), 190 E (glutamic acid), 226 Q (glutamine), and 227 Q (glutamine) for the receptor binding site. A comparison of the two strains of D1 and D7 with other strains in the gene bank showed that these isolates had identical sequences in the amino acid positions in the cleavage sites and receptor binding parts with A/Chicken/Iran/12VIR/6930/1998, A/ Chicken/Iran/ZMT-101/1998 (i.e., the isolates used in this study), A/Chicken/Iran/TH79/2000, and A/ Chicken/Saudi Arabia/CP7/1998 (Table 1). In the present study, the results of the nucleotide sequencing of the isolates showed a 99% similarity between the amino acids of these two isolates with the isolate of A/Chicken/Iran/ZMT-101/1998. In addition, the similarity of the studied HA gene with the isolated H9N2 virus from Saudi Arabia and Hong Kong in 1998 was reported as 99%.
Table 1.
Comparison of amino acid residues in two main sites of hemagglutinin genes
| Strain | Cleavage site | Receptor-binding site | ||||||
|---|---|---|---|---|---|---|---|---|
| 326 | 327 | 328 | 329 | 183 | 190 | 226 | 227 | |
| H3 | R * | Q | T | R | H | E | L | Q |
| A/Chicken/Iran/D1/2016 (H9N2) | R | S | S | R | H | E | Q | Q |
| A/Chicken/Iran/D7/2016 (H9N2) | R | S | S | R | H | E | Q | Q |
| A/Chicken/Iran/ZMT-101/1998 (H9N2) | R | S | S | R | H | E | Q | - |
| A/Chicken/Iran/12VIR/6930/1988 (H9N2) | - | - | - | - | - | - | - | - |
| A/Chicken/Iran/TH79/2000 (H9N2) | - | - | - | - | - | - | - | - |
| A/Chicken/Saudi Arabia/CP7/1998 (H9N2) | - | - | - | - | - | - | - | - |
| A/Chicken/Iran/av1221/1998 (H9N2) | - | - | - | - | - | A | - | - |
| A/Chicken/Iran/584/2000 (H9N2) | - | - | - | - | - | A | - | - |
| A/Chicken/Iran/11T/99 (H9N2) | - | - | - | - | - | A | - | - |
| A/Chicken/Pakistan/2/1999 (H9N2) | - | - | - | - | - | A | - | - |
| A/Chicken/Iran/661/1998 (H9N2) | - | - | - | - | - | A | L | - |
| A/Chicken/Iran/698/1998 (H9N2) | - | - | - | - | - | A | L | - |
| A/Chicken/Israel/90658/2000 (H9N2) | - | - | - | - | - | A | L | - |
| A/Chicken/Iran/TH77/1998 (H9N2) | - | - | - | - | - | E | L | - |
| A/Chicken/Iran/SS1/1998 (H9N2) | - | - | - | - | - | E | L | - |
| A/Parakeet/Chiba/1/97 (H9N2) | - | - | - | - | - | E | L | M |
| A/Chicken/Dubai/338/2001 (H9N2) | - | - | - | - | H | A | L | M |
| A/Chicken/Dubai/383/2002 (H9N2) | - | - | - | - | H | A | L | Q |
| A/Parakeet/Narita/92A/98 (H9N2) | - | - | - | - | - | E | M | Q |
| A/Chicken/Korea/25232-MS96CE6 /1996 (H9N2) | A | - | Y | - | - | - | Q | - |
| A/Turkey/Wisconsin/1/1966 (H9N2) | V | - | S | - | - | - | - | H |
| A/Swine/Hong Kong/9/98 (H9N2) | R | S | S | R | N | V | L | Q |
Amino acid abbreviations: R: Arginine; S: Serine; H: Histidine; A: Alanine; E: Glutamic acid; L: Leucine; Q: Glutamine; V: Valine; T: Threonine; M: Methionine
Phylogenetic tree analysis.
The phylogenetic tree of the HA gene under the H9N2 subtype with the sequence data of the two Iranian samples from the present study and 34 other isolates in the gene bank was drawn using the MEGA 7 software through maximum likelihood tree based on the Tamura-Nei model. The study of the phylogenetic tree, based on nucleotide sequence analysis, showed the highest similarity between D1 and D7. Clade 1 included the Iranian isolates isolated within 1998-2004, including A/Chicken/Iran/ZMT-101/1998 isolate shown by JX465626, two Iranian isolates shown in the present study by MF114292 and MF114293, and Saudi Arabia isolate shown by CY081264. In addition, they are placed in the same subcategory of clade 1. This clade indicated that the Saudi Arabian isolates of H9N2 subtypes of AIV had a close relationship with the Iranian isolates. Clade 2 included the isolates from the Middle East, Israel, Pakistan, United Arab Emirates, and Japan. In clade 3, there were six isolates from Iran which were examined within 2004-2005. The strains from Hong Kong (shown by KX879589), one human strain of influenza virus from the USA (Wisconsin 66; shown by CY087824), and one strain from Korea (shown by KF188345) are all placed in clade 4. This group was more distant from the other groups in terms of genetic status and position in the phylogenetic tree (Figure 3).
Figure 3.
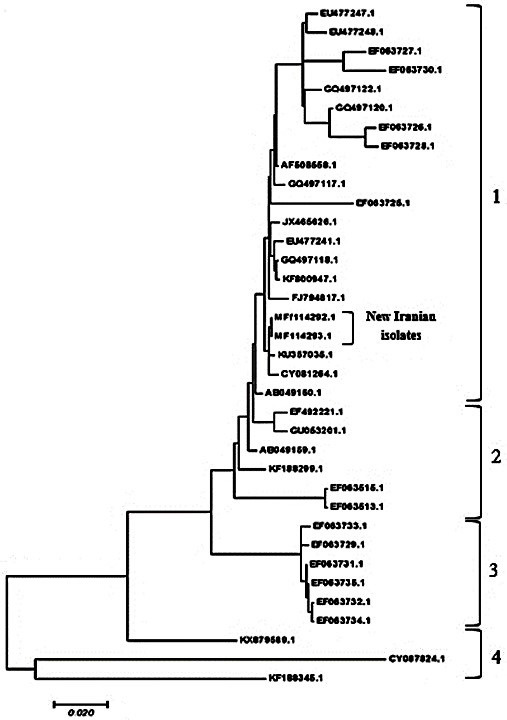
Phylogenetic tree related to the H9N2 subtype hemagglutinin gene of 2 new Iranian strains and 34 selected subtypes illustrating in four distinct clades; two strains of D1 and D7 highly related and belonging to a sister group
Discussion
The main objective of the present study was to determine the shedding of influenza virus (H9N2) in SPF chickens using the rRT-PCR molecular technique and examine gene changes in the cleavage site of the above-mentioned virus and compare them with other influenza viruses. Poultry influenza virus is circulating in many parts of the world, particularly in Iran, and chickens are susceptible to the virus ( Mehrabanpour et al., 2011 ). The H9N2 subtype virus was firstly detected and reported in the north of Iran in 1998 ( Pazani et al., 2008 ). With the constant circulation of the virus in various hosts, this type of virus was genetically modified and reported as spreading to vast areas of Asia, Europe, Middle East, and Africa ( Lee et al., 2007 ; Elbayoumi et al., 2013 ). In the present study, a number of viral shedding particles were compared by the rRT-PCR molecular method at various times after inoculation, with spreading pattern in the cloacal and oropharyngeal routes. The obtained results indicated that the type of infection and exposure of the virus in the host’s tissue played an important role in the manner of virus shedding. According to the obtained findings, virus shedding was at its highest in the oropharyngeal route on the 1st to 4th day after inoculation. Moreover, based on the investigation of the spread of the virus in the cloacal route, it was observed to be at its highest on the 2nd to 6th day after inoculation. The findings clearly indicated that the virus titer in the oropharyngeal route was higher than that reported for the cloacal route. The above-mentioned results are in line with the findings of previous studies because as Ge pointed out in his studies, LPAIVs replicate in the upper respiratory system and epithelial part of the intestine, in which there are trypsin-like enzymes. Cell surface receptors for AIV were glycoconjugate receptors containing sialic acid compounds. Consequently, AIV can bind with HA glycoprotein containing sialic acid; therefore, the main route of virus shedding is reported to be oropharyngeal ( Ge et al., 2012 ). The findings of a study carried out by Monne et al. (2008) indicated that the poultry viruses in Iran were closely related and likely to be of a common source, with the closest genetic affinity with the influenza viruses from India, Tunisia, Iraq, and United Arab Emirates ( Monne et al., 2008 ; Homayounimehr et al., 2010 ). Among LPAIVs, H9 subtypes attract more attention due to their direct transmission to humans and formation of new pandemics ( Cameron et al., 2000 ). According to studies conducted to date, the phylogenetic tree survey of the H9N2 virus showed that there are two branches of these viruses. The first and second branches belong to the American H9N2 viruses and European-Asian H9N2 viruses (Eurasian branch), respectively ( Lee et al., 2007 , Bashashati et al., 2013 ). Eurasian viruses are divided into three subcategories. The first subcategory includes the viruses isolated from Korea and Japan, and the indicator of this group is A/Chicken/ Korea/ MS96/96. The second subcategory consists of the strains isolated from humans in Hong Kong, whose indicator is A/The Quail/Hong Kong/G1/97. This type of virus also caused disease in the Middle East and Pakistan. The third branch of the virus was isolated from China (Beijing-like), whose representatives are A/Chicken/Beijing/1/94 and A/Duck/Hong Kong/ Y280/97 viruses ( Lee et al., 2007 ). In Iran, for the first time in 1998, the H9N2 subtype was isolated and identified as A/Chicken/Iran/259/1998 in the Research and Diagnosis Department of Poultry Diseases of Razi Vaccine and Serum Institute and College of Veterinary Medicine in University of Tehran ( Vasfi Marandi et al., 2002 ). The 1618-bp nucleotide sequencing of the 4th HA gene in the samples of the present study indicated the highest similarity (99%) with the recorded sequences of foreign samples, including CY081264 for Saudi Arabia (1998), AB049160 (Narita) for Hong Kong (1998), EF492221 for Israel (2000), and GU053201 for the Middle East (1999). However, the samples of the present study shown by MF114293 and MF114292 indicated the greatest difference in nucleotide sequences and least similarity with the foreign samples determined in the phylogenetic tree, including KX879589 for Hong Kong (1998), CY087824 for the American human strain (Wisconsin 66), and KF188345 for Korea (1996). The findings of the current study showed that the Iranian AIV is similar to the Eurasian H9N2 nonpathological viruses, and the lack of major genetic changes in the HA gene has not led to the emergence of a new pathotype. According to Cameron et al., the HA genes isolated from viruses in Germany, Pakistan, Saudi Arabia, and Iran within 1998-1999 are closely linked with the HA genes of human H9N2 and HK/G1/97 viruses isolated from Hong Kong ( Cameron et al., 2000 ). The results of studies conducted by Homayounimehr et al. (2010) indicated that the sequence of amino acids isolated from these studies underwent several changes. In the present study, there was no close relationship between the new D1 and D7 isolates with human H9N2 strains and HK/G1/97 strain. According to the sequences of amino acids shown in Table 5, the HA gene isolated from AIV in birds in ZMT-101, 12 VIR, CP7, 25232-MS96CE6, and Wisconsin strains at 183, 190, 226, and 227 positions have H- E - Q - Q amino acids, respectively. The present study also confirmed the similarity and uniformity of the sequences. The studies carried out by Homayounimehr et al. (2010) and Moosakhani et al. (2010) have raised the importance of the type of amino acid at position 190 for the binding tendency with sialic acid bound to galactose with α-2, 6 bind. In the aforementioned studies, it was stated that the binding tendency is at its highest when the amino acid type is V (valine). In addition, the moderate binding tendency happens at the presence of the T amino acid (threonine), and the weakest binding tendency happens at the presence of A amino acid (alanine) ( Homayounimehr et al., 2010 ). The present samples at position 190 are determined to be E amino acid (glutamic acid) (Table 1). The presence of L (leucine) amino acid at position 226 is a well-known sequence of H3 type and human influenza which is not observed in the isolates of birds. Accordingly, the D1 and D7 samples in the present study were believed to have the Q amino acid sequence (glutamine) (Table 1).
The specific receptors of HA play a decisive role in determining the domain of hosting influenza viruses. It has been observed that the AIVs detect SA α-2, 3 receptors for binding; nevertheless, human influenza viruses preferably choose SA α-2, 6 receptors. Therefore, the HA genes with a sequence of L amino acid (leucine) at position 226 have the tendency to bind with SA α-2,6; however, the genes with Q amino acid sequence (glutamine) will be attached to SA α-2,3 receptors ( Homayounimehr et al., 2010 ). In this regard, Wan and Perez suggested that when Q amino acid was replaced with L at position 226, the HA gene would make it possible for the H9N2 virus to infect nonsialic cells. Consequently, they can grow and have a greater effect on human epithelial cell culture; this raises the likelihood of an increase in the severity of infection in humans ( Wan et al., 2008 ; Homayounimehr et al., 2010 ). In previous studies investigating the sequences of a cut point in the HA gene, it was observed that the highly pathogenic viruses had an arginine amino acid (R) at position 328 with an R/K-X-R-R motif ( Wan et al., 2008 ; Homayounimehr et al., 2010 ). The findings of the present study showed that the D1 and D7 isolates at position 328 contained serine (S) amino acid, and they had glutamine (Q) amino acid at position 226 which could not bind with sialic acid and attach to galactose with the α-2, 6 bond. Therefore, the nonpathogenicity was preserved and subsequently had no pathogenic danger for humans. For a better perception, the nucleotide sequences of the HA gene in the isolates of A/Swine/Hong Kong/9/98 (H9N2) and H3 strain pathogens of humans should be considered, as listed in Table 1. These strains are pathogenic with (S and T) amino acids at position 328 and (L) amino acids at position 226. The data obtained from the present study together with those from the previous studies are very significant for the recognition of AIVs since some of these viruses may pose a threat to community health. For instance, some years ago, the H9N2 subtype virus was isolated from 8 individuals in Hong Kong with the symptoms similar to those of influenza. Therefore, although it is not severely pathogenic for the birds, the H9N2 is a virus that has the potential to provide the best gene combination for human infection and get passed through the mammals.
To sum up, given that the main route of the virus replication is in the areas in which trypsin-like enzymes are present, the oropharyngeal route in chickens was observed to be the main replication site for the influenza virus through measuring the high titer of the viral particles by the rRT-PCR molecular method. In phylogenetic studies, two D1 and D7 isolates from SPF chickens were compared with other isolates in the gene bank revealing that these isolates belonged to the Eurasian subcategory. The phylogenic analysis in the present study indicated that the D1 and D7 isolates had a similarity of 99%. The amino acids at 326, 327, 328, and 329 positions in the gene cleavage site of the Iranian ZMT-101 strain of the HA gene were identical; therefore, the sequence of these isolates was reported as R-S-S-R and similar to each other. This similarity and unchangeability in the HA gene in the isolates of the present study indicated that a new pathogen of the virus was not created. Therefore, they pose no pathogenic risk to humans. However, slight changes in the unnecessary areas of the HA gene warn that some of these viruses may be able to become super-acute viruses and have the potentiality of creating genetic rearrangements with other influenza viruses. Consequently, given the circulation of H9N2 viruses in the poultry flocks, exposed individuals, and changes toward human infection, it is essential to identify methods in order to fight against this virus.
Ethics
We hereby declare all ethical standards have been respected in preparation of the submitted article.
Grant Support
This study was supported by Razi Vaccine and Serum Research Institute of Shiraz, Iran, and Istituto Zooprofilattico Seprimentale delle Venezie in Italy.
Conflict of Interest:
The authors declare that they have no conflict of interest.
Acknowledgement
The authors would like to appreciate the cooperation of Dr. Giovanni Cattoli as well as Virology and Molecular Biology Department of Istituto Zooprofilattico Seprimentale delle Venezie.
References
- 1.Bashashati M, Vasfi Marandi M, Sabouri F. Genetic diversity of early (1998) and recent (2010) avian influenza H9N2 virus strains isolated from poultry in Iran. Arch Virol. 2013;158(10):2089–100. doi: 10.1007/s00705-013-1699-2. [DOI] [PubMed] [Google Scholar]
- 2.Cameron KR, Gregory V, Banks J, Brown IH, Alexander DJ, Hay AJ, Lin YP. H9N2 subtype influenza A viruses in poultry in pakistan are closely related to the H9N2 viruses responsible for human infection in Hong Kong. Virology. 2000;278(1):36–41. doi: 10.1006/viro.2000.0585. [DOI] [PubMed] [Google Scholar]
- 3.Costa-Hurtado M, Afonso CL, Miller PJ, Spackman E, Kapczynski DR, Swayne DE, Shepherd E, Smith D, Zsak A, Pantin-Jackwood M. Virus interference between H7N2 low pathogenic avian influenza virus and lentogenic Newcastle disease virus in experimental co-infections in chickens and turkeys. Vet Res. 2014;45(1):1–11. doi: 10.1186/1297-9716-45-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Elbayoumi KM, Mahgoub K, Mekky H, Hassan ER, Girh ZMS, Maatouq A, et al. Molecular detection of H5N1, H9N2 and Newcastle disease viruses isolated from chicken in mixed infection in Egypt. World Appl Sci J. 2013; 27: 44–50. [Google Scholar]
- 5.Fouchier RA, Munster V, Wallensten A, Bestebroer TM, Herfst S, Smith D, Rimmelzwaan GF, Olsen B, Osterhaus AD. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J Virol. 2005;79(5):2814–22. doi: 10.1128/JVI.79.5.2814-2822.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ge S, Zheng D, Zhao Y, Liu H, Liu W, Sun Q, Li J, Yu S, Zuo Y, Han X, Li L, Lv Y, Wang Y, Liu X, Wang Z. Evaluating viral interference between Influenza virus and Newcastle disease virus using real-time reverse transcription-polymerase chain reaction in chicken eggs. Virol J. 2012 1;9:128. doi: 10.1186/1743-422X-9-128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Homayounimehr AR, Dadras H, Shoushtari A, Pourbakhsh SA. Sequence and phylogenetic analysis of the haemagglutinin genes of H9N2 avian influenza viruses isolated from commercial chickens in Iran. Trop Anim Health Prod. 2010;42(6):1291–7. doi: 10.1007/s11250-010-9565-7. [DOI] [PubMed] [Google Scholar]
- 8.Hosseini H, Ghalyanchilangeroudi A, Fallah Mehrabadi MH, Sediqian MS, Shayeganmehr A, Ghafouri SA, Maghsoudloo H, Abdollahi H, Farahani RK. Phylogenetic analysis of H9N2 avian influenza viruses in Afghanistan (2016-2017) Arch Virol. 2017;162(10):3161–3165. doi: 10.1007/s00705-017-3474-2. [DOI] [PubMed] [Google Scholar]
- 9.Karimi S, Ghalyanchilangeroudi A, Madadgar O, Najafi H, Maghsoudlo H. Molecular characterization and phylogenetic study based on matrix gene of avian influenza viruses (H9N2) in Iran during 1998-2008. J Vet Res. 2015;70: 147–153. [Google Scholar]
- 10.Lee YJ, Shin JY, Song MS, Lee YM, Choi JG, Lee EK, Jeong OM, Sung HW, Kim JH, Kwon YK, Kwon JH, Kim CJ, Webby RJ, Webster RG, Choi YK. Continuing evolution of H9 influenza viruses in Korean poultry. Virology. 2007;359(2):313–23. doi: 10.1016/j.virol.2006.09.025. [DOI] [PubMed] [Google Scholar]
- 11.Liolios L, Jenney A, Spelman D, Kotsimbos T, Catton M, Wesselingh S. Comparison of a multiplex reverse transcription-PCR-enzyme hybridization assay with conventional viral culture and immunofluorescence techniques for the detection of seven viral respiratory pathogens. J Clin Microbiol. 2001;39(8):2779–83. doi: 10.1128/JCM.39.8.2779-2783.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Matrosovich M, Tuzikov A, Bovin N, Gambaryan A, Klimov A, Castrucci MR, Donatelli I, Kawaoka Y. Early alterations of the receptor-binding properties of H1, H2, and H3 avian influenza virus hemagglutinins after their introduction into mammals. J Virol. 2000;74(18):8502–12. doi: 10.1128/jvi.74.18.8502-8512.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mehrabanpour MJ, Fazel PD, Rahimian A, Hosseini MH, Moein H, Shayanfar MA. Newcastle Disease and Avian Influenza A Virus in Migratory Birds in Wetland of Boushehr-Iran. J Anim Vet Adv. 2011; 3: 229–234. [Google Scholar]
- 14.Monne I, Ormelli S, Salviato A, De Battisti C, Bettini F, Salomoni A, Drago A, Zecchin B, Capua I, Cattoli G. Development and validation of a one-step real-time PCR assay for simultaneous detection of subtype H5, H7, and H9 avian influenza viruses. J Clin Microbiol. 2008;46(5):1769–73. doi: 10.1128/JCM.02204-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Moosakhani F, Shoshtari AH, Pourbakhsh SA, Keyvanfar H, Ghorbani A. Phylogenetic analysis of the hemagglutinin genes of 12 H9N2 influenza viruses isolated from chickens in Iran from 2003 to 2005. Avian Dis. 2010;54(2):870–4. doi: 10.1637/9103-101309-Reg.1. [DOI] [PubMed] [Google Scholar]
- 16.Pazani J, Vasfi Marandi M, Ashrafihelan J, Marjanmehr S, Farid G. Pathological Studies of A / Chicken / Tehran / ZMT - 173/99 (H9N2) Influenza Virus in Commercial Broiler Chickens of Iran. Int J Poultry Sci. 2008; 7: 502–510. [Google Scholar]
- 17.Spackman E, Senne DA, Myers TJ, Bulaga LL, Garber LP, Perdue ML, Lohman K, Daum LT, Suarez DL. Development of a real-time reverse transcriptase PCR assay for type A influenza virus and the avian H5 and H7 hemagglutinin subtypes. J Clin Microbiol. 2002;40(9):3256–60. doi: 10.1128/JCM.40.9.3256-3260.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–9. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Toroghi R, Momayez R. Biological and molecular characterization of Avian influenza virus (H9N2) isolates from Iran. Acta Virol. 2006;50(3):163–8. [PubMed] [Google Scholar]
- 20.Vasfi Marandi M, Bozorg Mehrifard MH, Tabatabaei SM. A Seroepidemiologic study of avian influenza (H9N2) in Iran. Sci Res Iran Vet J. 2002; 8: 23–31. [Google Scholar]
- 21.Wan XF, Nguyen T, Davis CT, Smith CB, Zhao ZM, Carrel M, et al. Evolution of highly pathogenic H5N1 avian influenza viruses in Vietnam between 2001 and 2007. PloS one. 2008; 3: e3462. doi: 10.1371/journal.pone.0003462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhu G, Wang R, Xuan F, Daszak P, Anthony SJ, Zhang S, Zhang L, He G. Characterization of recombinant H9N2 influenza viruses isolated from wild ducks in China. Vet Microbiol. 2013;166(3-4):327–36. doi: 10.1016/j.vetmic.2013.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]