
Figure 6.
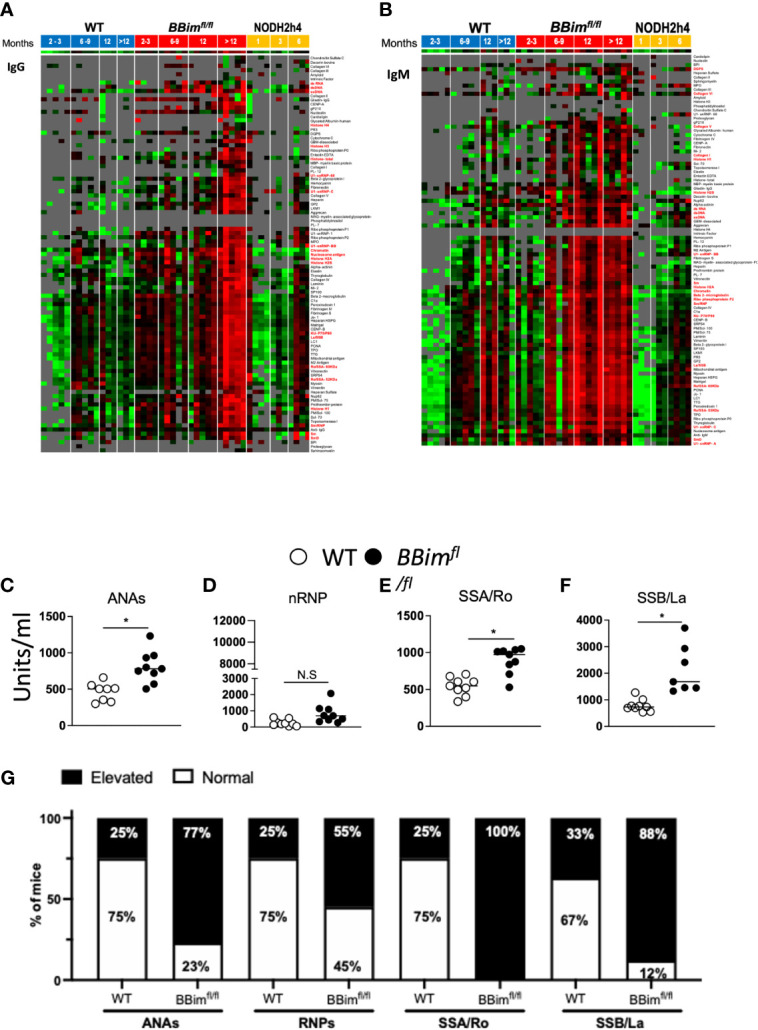
BBimfl/fl mice exhibit elevated titers of SLE/SS signature autoAbs. Serum from mice of indicated age groups were screened for autoAbs by hybridizing to an array containing over 90 autoantigens. Each serum specimen was taken from a different mouse so each time point would be independent. Red boxes indicate greater reactivity than the average for each antigen and green boxes indicate lower reactivity than the average for each antigen. Reactivities close to the mean are displayed in black/gray. Some of the SS and SLE-associated autoAb names are highlighted in red. (A, B) Heat maps with clustering of WT (left), BBimfl/fl (middle) and NODH2h4 (right). Supervised clustering of autoAbs was performed with normalized signal intensities for baseline IgG and IgM autoAbs. (A) IgG and (B) IgM autoAbs showing higher reactivity (red) appeared in serum of BBimfl/fl mice by 6-9 months. At 12 months, additional IgM antibodies emerged and persisted, which were again followed by the subsequent development of IgG antibodies in most instances. In each case tested (with the exception of PR3 antibodies (vasculitis-associated) which remained persistently IgM only), IgM antibodies appeared at time points either concurrent with or prior to the emergence of IgG antibodies, and both IgM and IgG persisted through all subsequent time points. The autoantibodies that emerged included all of the tested SS-associated antibodies (Ro52/SS-A, Ro60/SS-A, La/SS-B, CENP-B), and a variety of lupus-associated autoantibodies (chromatin, ssDNA, alpha-actinin, vitronectin, snRNP, Beta2-GPI, PCNA, nucleosome Ag, Sm-D, Histone H2B, peroxiredoxin 1, ribophosphoprotein P0, myosin, Heparan HSPG, Matrigel, Vitronectin, Heparin, collagen IV). Additional autoantibodies emerging in the same time frame included those targeting some autoimmune hepatitis antigens (LKM1, mitochondrial Ag, LC-1), some myositis-associated antigens (SRP54, Jo-1, Nup6.2), celiac disease –associated targets (DGPS, TTG), as well as the following antigens: C1alpha, the Crohn’s-associated antigen GP2, and the thyroiditis-associated antigen TPO. At 12 months, additional IgM antibodies emerged and persisted, followed by the development of IgG in most instances includingU1-snRNP-BB, U1-RNP-C, laminin, hemocyanin, aggrecan, fibrinogen s, autoimmune hepatitis antigens (M2 antigen, SP100), and a scleroderma/myositis-associated target (PM-Scl75). However, some of IgM autoAbs were not accompanied/followed by IgG autoAbs, exemplified by the myositis –associated antigen PL12 and neuropathy-associated myelin associated glycoprotein-Fc (MAG) and collagen V (in older mice).In addition, aged BBimfl/fl mice developed some additional IgM autoAbs to lupus-associated autoantigens in the majority of tested mice including GBM-associated, and prothrombin protein, which were associated with concurrent emergence of IgG against the same targets. Several autoAbs were upregulated only in aged mice and only as IgG, including additional lupus-associated specificities (U1-snRNP68, U1-snRNP1, Sm, total histone, histone H2A, Histone H1, Heparan Sulfate, Entactin-EDTA, collagen II, fibronectin, elastin, fibrinogen IV, ribophosphoprotein P1, ribophosphoprotein p2), a myositis-associated target (Mi-2), some scleroderma-associated specificities (CENP A, Ku P70-p80, Scl-70, Topoisomerase I, PM-Scl100), an autoimmune hepatitis-associated target (gp210), a thyroiditis-associated antigen (thyroglobulin), and the vasculitis-associated target MPO. (C–F) Serum fromBBimfl/fl and WT mice (age ≥ 24 weeks old) were assessed for autoAbs distinctive for SS and SLE by ELISA. (G) Representative bar graphs show the frequency of elevated (filled bar) and normal (open bar) autoantigens titers in WT and BBimfl/fl mice. Elevated titers were determined by calculating the positive index as stated in the ELISA kit protocol (alpha diagnostic international). Data is representative of >3 independent experiments. Not significant (N.S) P > 0.05, *P ≤ 0.05, calculated by Students T-test and non-parametric Mann-Whitney test.