
Figure 4.
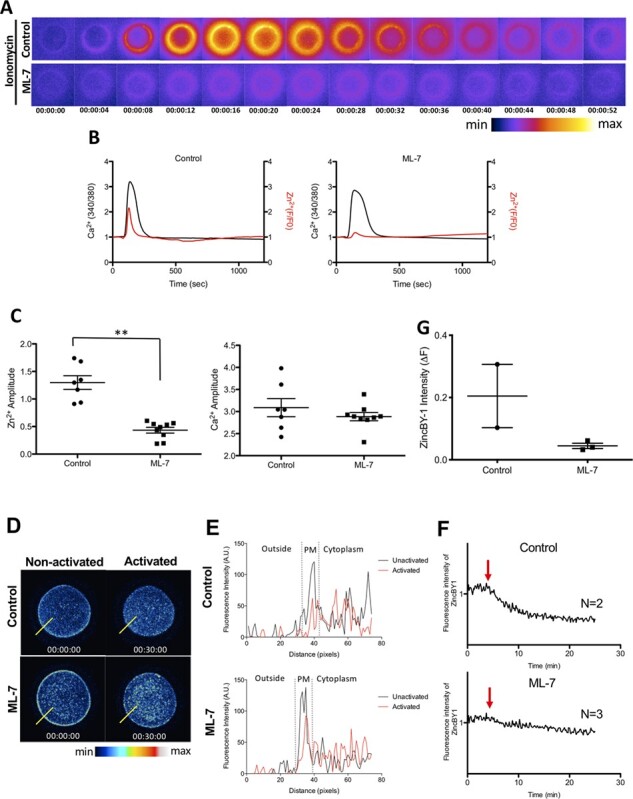
The zinc spark is blunted, and labile zinc localisation to the cortex is reduced, in ML-7-treated human eggs after activation by ionomycin. (A) A time-lapse montage demonstrating representative zinc sparks (detected by 20 μM Fluozin-3) in control and ML-7-treated human MII eggs in response to activation by 5 μM ionomycin. The LUT colour scale is displayed at the bottom. (B) Representative time traces of normalised zinc fluorescence (red, F/F0) and calcium fluorescence (black, 340/380) are shown for control and ML-7-treated human MII eggs. (C) A dot graph quantifying the amplitudes of the zinc sparks and calcium transients in control and ML-7-treated eggs; ML-7-treated eggs demonstrated a 67% reduction in their zinc spark amplitude compared with controls (** P < 0.001; non-treated activated eggs: 1.298 ± 0.1467, n = 7; vs. ML-7-treated activated eggs: 0.4336 ± 0.0589, n = 9). (D) Representative images of ZincBY-1 staining in human eggs, demonstrating their intracellular zinc intensity and location between control and ML-7-treated eggs, both before and after activation by ionomycin. The LUT colour scale is displayed at the bottom. (E) A representative histogram of ZincBY1 fluorescence intensity across the PM of human eggs after activation in control versus ML-7 treatment groups (assembled using the yellow lines drawn through eggs in D). (F) Representative time traces of normalised labile zinc fluorescence in the cortical region of activated human eggs. Red arrows signify the addition time of ionomycin. (G) A dot graph quantifying the change in ZincBY-1 intensity (ΔF) across activation. Data were presented as the median. (* P < 0.05, ** P < 0.001).