

ABSTRACT
Butterflies and moths (Lepidoptera) are one of the most studied, diverse, and widespread animal groups, making them an ideal model for climate change research. They are a particularly informative model for studying the effects of climate change on species ecology because they are ectotherms that thermoregulate with a suite of physiological, behavioural, and phenotypic traits. While some species have been negatively impacted by climatic disturbances, others have prospered, largely in accordance with their diversity in life‐history traits. Here we take advantage of a large repertoire of studies on butterflies and moths to provide a review of the many ways in which climate change is impacting insects, animals, and ecosystems. By studying these climate‐based impacts on ecological processes of Lepidoptera, we propose appropriate strategies for species conservation and habitat management broadly across animals.
Keywords: asynchrony, butterfly, insect, morphology, moth, parasitoid, phenology, range shift, trophic mismatch, voltinism
I. INTRODUCTION
Climate change is affecting ecosystems on all scales from individual genotypes to entire communities (Wilson & Maclean, 2011; Wymore et al., 2011; Scheffers et al., 2016; Cohen, Lajeunesse & Rohr, 2018). Despite a substantial body of literature, it is still difficult to predict ecological responses to climate change. Evidence is scattered across taxa, habitats, populations, and communities (Parmesan & Yohe, 2003; Parmesan, 2006; Merilä & Hendry, 2014; Lister & Garcia, 2018; Wepprich et al., 2019), making it difficult to discern broad patterns in responses across time and space. Scheffers et al. (2016) provided a unifying synthesis of evidence on climate change impacts across the hierarchy of biological organization but to do so, the authors had to collate evidence from diverse taxa and ecological systems. Here we complement this approach with a deeper synthesis of a single, well‐studied taxon to recommend best conservation and management practices for organisms with overlapping traits and ecologies.
Insects have been proposed as informative models for testing climate change impacts on ecological systems because they show rapid responses to changes in their environment due to short generation times and sensitive ecological requirements (Nadeau, Urban & Bridle 2017). There is mounting evidence that they are disappearing rapidly, with climate change being a major contributing factor (Oliver et al., 2015; Forister, Pelton & Black, 2019; Soroye, Newbold & Kerr, 2020; Wagner, 2020; Halsch et al., 2021). Understanding the effects of climate change on insects is therefore a top priority for conservationists, as they play an important role in shaping Earth's biota (Misof et al., 2014), and make up the largest proportion of animal diversity and biomass (Bar‐On, Phillips & Milo, 2018). Unfortunately, the life histories of many insect groups are not well understood. The one exception is the butterflies and moths (Lepidoptera), undoubtedly the best‐studied insect lineage, and is already responding to climate change, habitat loss, and habitat fragmentation (Warren et al., 2001; Wilson & Maclean, 2011; Fox, 2013; Belitz et al., 2018; Maurer et al., 2018; Ellis et al., 2019).
Butterflies and moths are well suited as a model for uncovering patterns in the effects of climate change on ecosystems since the literature on this group demonstrates how organismal (genetics, physiology, behaviour, morphology), phenological (host synchrony, voltinism), population‐level (geographic ranges), and community‐level (trophic interactions e.g. parasitoid–herbivore) processes interact and respond to change. They are an ideal group for investigating associations between species traits and range shifts due to the availability of accurate scientific information on their biology, and the distributional data from citizen science monitoring schemes. Lepidoptera have proved useful in monitoring ecological and evolutionary responses to climate because they capture the diversity of responses across insects: (i) they possess a suite of genetic, physiological, and morphological traits with known sensitivity to climatic variables. Several studies have recorded their capacity for rapid evolutionary change via genetic and phenotypic responses to current warming. (ii) They have complex life cycles that span a wide array of life‐history strategies relating to development time, emergence time, foraging behaviour, survivorship, and voltinism. (iii) They exemplify the complexity of geographical responses to climate change – some species of Lepidoptera have undergone local extinctions, while others have experienced population increases.
To complement Scheffers et al. (2016), who summarized evidence across multiple taxa and systems, we focus on the extensively studied butterflies and moths to uncover systematic biological responses to climate change within a phylogenetically related but diverse group of organisms in a wide range of habitats and ecosystems. We use case studies from contemporary literature to identify and discuss climate change impacts on ecological processes (Fig. 1), determine research gaps, and propose future directions for climate change research. Our synthesis on a well‐studied model taxon serves to illustrate many of the general trends purported in the ecological climate change literature.
Fig 1.
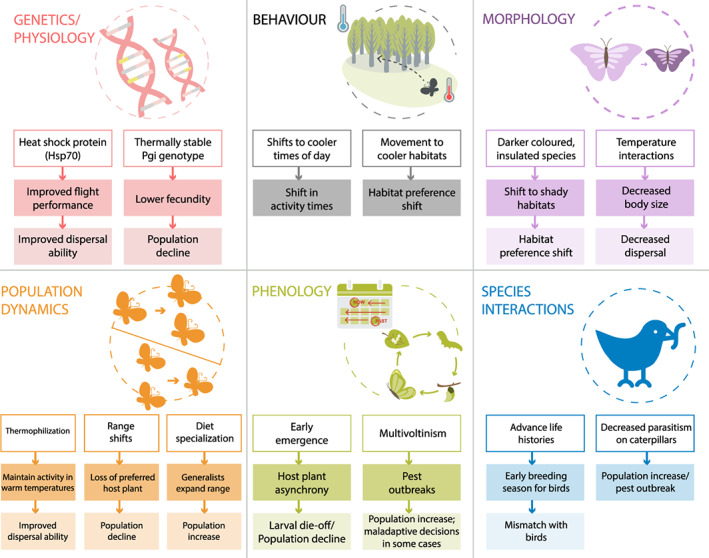
Examples of predicted and observed responses of Lepidoptera to climate change. Each response within the ecological processes has at least one supporting case study. Case studies include: Dewar & Watt (1992); Williams & Liebhold (1995); Parmesan et al. (1999); McLaughlin et al. (2002); Crozier (2003); Parmesan & Yohe (2003); Visser et al. (2004); Stireman et al. (2005); Wilson et al. (2005); Thomas (2005); Kiritani (2006); Parmesan (2006); Nogués‐Bravo et al. (2007); Hellmann et al. (2008); Menéndez et al. (2008); Schweiger et al. (2008); Dukes et al. (2009); Pelini et al. (2009); Altermatt (2010a ); Singer & Parmesan (2010); Chen et al. (2011); Finkbeiner et al. (2011); Betzholtz et al. (2012); Breed et al. (2013); Bonebrake et al. (2014); Kingsolver & Buckley (2014); Aalberg Haugen & Gotthard (2015); Duque et al. (2015); Filz & Schmitt (2015); Van Dyck et al. (2015); Chuang & Peterson (2016); Kleckova & Klecka (2016); Melero et al. (2016); Molina‐Martínez et al. (2016). Pgi, phosphoglucose isomerase.
II. ORGANISMAL RESPONSES
The most obvious impact of climate change is that species adapted to cooler environments will find it increasingly difficult to acclimate to warmer ones. In such circumstances, they may escape extinction by either adapting in situ to the warmer climate or moving to a cooler one, depending on the respective availability of genotypes that are heat‐tolerant and/or predisposed to high dispersal. In this section, we discuss how these adaptive genotypes respond to climate change in Lepidoptera.
(1). Genetic
Genetic variation can allow a species to tolerate different environments when selection acts on dispersal capacity (Canale & Henry, 2010). The effects of anthropogenic climate change have already altered the selective pressures on butterfly populations, leading to allele frequency shifts associated with dispersal (Karl, Schmitt & Fischer, 2009; Kleckova & Klecka, 2016). Genetic polymorphisms in the Pgi gene encoding phosphoglucose isomerase have been shown to influence several key life‐history traits in adult insects, including dispersal, flight metabolism, longevity, and fecundity (Watt, 1983; Saastamoinen, Ikonen & Hanski, 2009; Kallioniemi & Hanski, 2011). In butterflies, Pgi heterozygotes have increased fitness in cooler climates as they are capable of flying at lower ambient temperatures than their homozygote counterparts (Niitepõld et al., 2009). Genetic variation in Pgi in the Glanville fritillary (Melitaea cinxia) directly affects dispersal rates and metapopulation dynamics (Haag et al., 2005; Hanski & Saccheri, 2006; Saastamoinen & Hanski, 2008). Similarly, in the map butterfly (Araschnia levana), individuals have higher levels of dispersive Pgi alleles at newly colonized sites, even though these individuals appear to have no morphological changes that improve flight performance such as increased wing or thorax size (Mitikka & Hanski, 2010). These results suggest that range expansions may be assisted by, or even dependent on, the selection of dispersive Pgi alleles in thermally challenged species.
Pgi alleles are also associated with heat resistance in Lepidoptera, but selection of resistant genotypes in warmer environments may be impeded by trade‐offs with other important life‐history traits. In Colias butterflies, the genotypes that are the most heat stable are also the least fecund, so selection for heat tolerance could greatly reduce population sizes (Watt, 1992). In general, Pgi genotypes that are associated with a narrow window of flight are more metabolically active, more fecund, and less heat resistant, whereas genotypes associated with a broad flight window have lower kinetic power, lower fecundity, and increased resistance to heat (Schneider & Root, 2002). Selection for heat‐resistant Pgi alleles under global warming may therefore be hampered by associated costs of decreased fecundity and reduced dispersal; however, Pgi is not directly influential in heat resistance in Lycaena tityrus (Fischer & Karl, 2010), so trade‐offs may vary among taxa. Future studies on the role of Pgi could be extremely important for conservation genetics, since it can be used to detect local adaptation and dispersal in changing environments (Wheat, 2010).
Another molecular marker that is likely to be important in the response to climate change is the heat shock protein (Hsp70). Hsp70 genes play a critical role in helping insects survive exposure to extreme temperatures by increasing heat tolerance (Wang et al., 2015). Under normal conditions, Hsp70 is expressed at very low levels, but expression increases quickly in response to extreme temperature‐related stress (Bahar et al., 2013). This response was observed in wide‐ranging moth pests, including the diamondback moth (Plutella xylostella) and setaceous Hebrew character moth (Xestia c‐nigrum) where the expression of several Hsp70 genes became more frequent when exposed to heat or cold stress (Bahar et al., 2013; Yang et al., 2015). Thus, sensitivity in gene expression of Hsp70 can be used to evaluate a species' ability to adjust to short‐term extreme temperatures (Bahar et al., 2013). Hsp70 genotypes are also associated with body temperature and metabolic rate prior to flight in M. cinxia (Mattila, 2015), which might allow improved flight performance under higher temperatures in the future. Taken together, Pgi and Hsp70 offer a robust comparison of key genes and phenotypes that are directly impacted by a changing climate. Although further genetic markers have been uncovered in other animals and plants (Hoffmann & Daborn, 2007; Franks, Weber & Aitken, 2014), markers need to be established for species from broader phylogenetic and ecological contexts (Varshney et al., 2018). The utility of Pgi and Hsp70 suggests that genes associated with thermoregulation and dispersal may be highly informative and would be a useful target in future research.
(2). Behavioural
Behavioural plasticity is especially important for ectotherms like butterflies and moths which must seek out the correct environment to keep their body temperature at an optimum (behavioural adaptations). For example, species living in alpine habitats hold their wings perpendicular to the sun (basking) while at rest to increase exposure to heat (Kingsolver, 1985). Conversely, those in warmer climates adopt a heat‐avoidance posture, like Chlosyne lacinia and Colias butterflies, which inhabit open habitats and orient away from the sun to avoid reaching lethal body temperatures (Bonebrake et al., 2014; Kingsolver & Buckley, 2014). In response to increasing temperatures and sunlight, heat avoidance and basking behaviours are likely to become more and less frequent, respectively. Species that exhibit avoidance behaviours could have a selective advantage in open, exposed environments where direct sunlight may push individuals towards their lethal limits (Bonebrake et al., 2014) but may also face context‐dependent challenges if posturing impacts camouflage or foraging time.
Seeking optimal microhabitats is likely to be the most common behavioural response of butterflies and moths to climate change. Species residing in cool habitats may take advantage of warmer microclimates to enhance their body temperature, while species in warmer habitats, such as those at lower elevations, may retreat to cooler, more shaded microhabitats (Kleckova & Klecka, 2016). For instance, larvae of the two swallowtail butterflies, the Apollo butterfly (Parnassius apollo) and pipevine swallowtail (Battus philenor) move between microhabitats depending on ambient temperature and sunlight (Nice & Fordyce, 2006; Ashton, Gutiérrez & Wilson, 2009). P. apollo larvae spend time thermoregulating in areas with reduced vegetation cover at higher, cooler elevations, whereas at lower, warmer elevations, they spend more time in sheltered, shady areas (Ashton et al., 2009). Caterpillars of the small eggar (Eriogaster lanestris) can reach their thermal optimum over a wide range of ambient temperatures by changing position within their layered, silken tents (Ruf & Fiedler, 2002). These examples suggest that climate warming may trigger widespread changes in microhabitat preference and increased microhabitat‐seeking behaviours. Species that exhibit a greater capacity for thermoregulatory behaviour may be the best suited to meet changes in temperature and solar radiation (cloud cover). Furthermore, the success of optimal habitat‐seeking will depend on other factors, such as the existing thermal requirements of a species, and whether optimal microhabitats are available (Kleckova & Klecka, 2016).
Sociality is another trait that has been associated with a species' tolerance to external temperatures, suggesting that social behaviours may become a selective advantage as global temperatures continue to change. A comparative study between two gregarious and two solitary moth species showed that gregarious larvae had a greater ability to adjust their body temperatures, while solitary species were more dependent on external temperatures (Bryant, Thomas & Bale, 2000). The thermoregulatory behaviour of tent caterpillars may also be linked to their gregarious nature (Ruf & Fiedler, 2002). With only a few examples, it is difficult to suggest whether this is a general trend, but this is an important area for future research.
The consequences of climate change will also depend on how well thermoregulatory behaviours synchronize with changing seasonal activity and reproduction, and how shade availability is affected by changes in vegetation cover (Kearney, Shine & Porter, 2009). Increased temperatures in certain environments may limit activity time and increase maintenance energy costs, such as those associated with seeking optimal microhabitats (Kearney et al., 2009). Additionally, microclimate has impacts on the phenology of host plants and nectar sources, which can in turn affect behaviour, such as in Edith's checkerspot (Euphydryas editha) caterpillars which disperse towards optimal microhabitats and host plants (Weiss, Murphy & White, 1988). These examples highlight the importance of incorporating information on behaviours such as microhabitat and host seeking into life‐history studies. Ecological characterizations are still sorely lacking across the tree of life, with 88–95% of eukaryotes still undescribed (Mora et al., 2011; Hawksworth & Lücking, 2017; Stork, 2018). Habitat and temperature requirements of under‐represented taxa, and the behavioural mechanisms used to maintain those requirements, are critical research areas to inform conservation strategies under climate change.
(3). Morphological
Morphological responses to climate change are some of the best documented, largely because of specimen records in natural history collections (e.g. MacLean et al., 2018). Several studies have demonstrated how decreased melanism and variable colour patterns may provide selective advantages in rising temperatures (Forsman, Betzholtz & Franzén, 2016; Scriber, 2020). Darker, more melanic wings allow butterflies to absorb more heat in cooler microclimates, and longer setae on the thorax help to retain heat and increase body temperatures at higher elevations (Kingsolver & Moffat, 1982). Phenotypic plasticity of these traits provides an adaptive route for a species to thermoregulate according to changing temperatures. For example, Colias meadii butterflies subjected to colder temperatures during pupal development have increased wing melanism and longer setae at high elevations (MacLean, Kingsolver & Buckley, 2016). Plasticity in larval colour pattern can also provide adaptive resilience. In the pipevine swallowtail (B. philenor), a greater proportion of red larvae are produced at warmer temperatures, and a black phenotype is maintained at cooler temperatures, allowing them to cope better with critical thermal extremes (Nice & Fordyce, 2006). For some European butterflies, dark‐coloured and lighter‐coloured individuals are selected in cooler and warmer climates, respectively (Zeuss et al., 2014). When temperatures are high, light‐coloured species can be active for longer periods during the day than dark‐coloured species, which broadens the range of suitable habitats at low latitudes. A similar trend is found in Australian butterflies but at much smaller spatial scales. Here, darker and larger butterflies preferred cool, shady habitats since they absorb heat faster compared to lighter‐coloured and smaller butterflies (Xing et al., 2016). Thus, climate warming may present new fitness costs or trigger habitat shifts, with darker coloured species shifting their behaviour and preference towards more shady habitats, shorter activity times and cooler regions, which might represent a last‐resort climate change refuge, after which localized extinctions of dark species might occur.
Near‐infrared reflectivity of some butterfly wings helps to control heat gain and may be selected for under climate change as temperatures increase (Munro et al., 2019). In general, heat‐avoidance morphologies such as reflectivity and reduced melanism will present an advantage for butterflies in warming climates, while plasticity or high intraspecific variation of any thermoregulatory morphology is likely to provide resilience in fluctuating environments prone to extreme weather events, which are predicted to occur more frequently under climate change. For example, species with more variable colour patterns may have an advantage over monotypic species (Forsman et al., 2016). However, some trade‐offs can be expected for traits like melanism, which also confer functions associated with immune responses or life histories (Clusella‐Trullas & Nielsen, 2020).
Intuitively, one might expect that largely nocturnal species may not benefit from either behavioural thermoregulatory modifications or melanism in response to climate exposure. Yet, Xing et al. (2018) show that geometrid moths, which are primarily nocturnal in activity, show disproportionately darker moth species at high elevations. Their result suggests that the distribution of colour lightness of nocturnal moths is affected by temperature and solar radiation, especially along climatically harsh environmental gradients where organisms are under strong selective pressures. An interesting topic for future research would therefore be to investigate the importance of dark colour patterns for the thermoregulatory activity and UV protection of night‐flying moths.
The effect of climate change on lepidopteran body size is more difficult to predict, with studies showing differing responses depending on phenology and existing plasticity. Across ectotherm taxa, warming global temperatures often appear to result in smaller body size (Ohlberger, 2013; Coulthard et al., 2019) by shortening development time (Sheridan & Bickford, 2011). Larger bodies have also been shown to reduce the time needed to raise body temperature for flight activity in cold environments (Nève & Després, 2020), an adaptation that may not confer the same advantages with increasing temperatures. In the Arctic butterflies Boloria chariclea and Colias hecla, decreases in body and wing size were observed in response to 17 years of rising summer temperatures in Greenland (Bowden et al., 2015). However, recent work suggests that the effect of temperature on body size in insects can be more complex and varied. For example, increased wing length is correlated with warmer temperatures during specific pupal periods in Anthocharis cardamines butterflies (Davies, 2019). The impact of temperatures experienced during development on adult body size in butterflies has also been analysed in museum specimens of Hesperia comma (Fenberg et al., 2016), Polyommatus argus, Polyommatus bellargus and Polyommatus coridon (Wilson, Brooks & Fenberg, 2019). The direction (increased or decreased wing length) of the response to temperature varied with developmental stage, sex, and generation, suggesting that complex ecological and evolutionary feedbacks within populations may be triggered by climate change.
Climate change, and warming temperature especially, can cause conflict between morphological traits like body and wing size, and ecological traits like dispersal. In cases where changing climates cause decreases in body size, a major consequence is reduced dispersal capacity and fecundity. Morphometric data from the swallowtail butterfly (Papilio machaon britannicus) show that, as populations contracted over time in England, selection against dispersal from isolated habitats led to morphological changes associated with decreased body size (Dempster, King & Lakhani, 1976). The converse of this process, whereby selection for reduced body size in warmer environments leads to associated loss of dispersal capacity, may therefore be expected under climate change. In Bornean geometrid moths, a reduction in wing size at high altitudes was tied to uphill range shifts of smaller species after four decades of warming (Wu et al., 2019); unequal redistribution of different sized species under climate change may therefore affect community size composition. In other cases, the effect on dispersal may be sex specific. The speckled wood butterfly (Pararge aegeria) generally has populations with larger adults and increased thorax size at newly colonized sites, but these phenotypic and evolutionary changes are more apparent in females than males which may impact responses to climate change (Hill, Thomas & Blakeley, 1999a ; Hughes, Dytham & Hill, 2007). There is evidence that dispersal ability is not always associated with changes in body and wing size. Finnish populations of M. cinxia have increased dispersal ability in newly colonized sites, but do not exhibit any changes in flight morphology (Hanski et al., 2002). However, Kuussaari, Nieminen & Hanski (1996) found that migratory female M. cinxia were larger (as measured by forewing length) than non‐migratory individuals, and Davies & Saccheri (2013) found that smaller (wing‐length) male A. cardamines were less dispersive in a localized English population than larger ones. Hence, the traits associated with dispersal may be cryptic (genetic, physiological, behavioural) or non‐cryptic (morphological), and these may vary among populations and species. Because high dispersal rates are a distinguishing feature of species living in rapidly changing environments (Hanski, Saastamoinen & Ovaskainen, 2006), future research focusing on dispersal‐linked morphological traits should yield promising results on range boundary expansion and colonization of novel habitats.
III. POPULATION‐LEVEL RESPONSES
(1). Population and range expansion
The rate at which individual species shift their range is dependent upon multiple internal traits and external drivers. Range redistributions in butterflies and moths, such as those observed across Europe, Asia, and North America, have been associated with ecological generalization, dispersal ability and reproductive rate (Crozier, 2003; Kiritani, 2006; Parmesan, 2006; Dukes et al., 2009; Pöyry et al., 2009; Forister et al., 2010; Breed, Stichter & Crone, 2013). For example, the sachem (Atalopedes campestris) is a coastal skipper butterfly with tropical origins formerly confined to the southwestern USA due to winter temperatures. Over the past 40 years, A. campestris has expanded its range northward up the west coast in response to warmer winters (Crozier, 2003). This species has been able to expand its range because it is not habitat or dispersal limited (Crozier, 2003). Thermophilization of communities is expected to occur across temperate, boreal and arctic ecosystems as a result of warm‐adapted species being favoured under climate change (Gottfried et al., 2012; Duque, Stevenson & Feeley, 2015). Shifting ranges have already been observed for species in several butterfly families: Hesperiidae (propertius duskywing Erynnis propertius), Lycaenidae (brown argus Aricia agestis), Nymphalidae (speckled wood butterfly P. aegeria), and Papilionidae (giant swallowtail Papilio cresphontes, anise swallowtail P. zelicaon) (Hellmann et al., 2008; Menéndez et al., 2008; Finkbeiner et al., 2011; Breed et al., 2013), as well as agricultural and forest moth pest species like the African maize stalk borer (Busseola fusca) and the pine processionary moth (Thaumetopoea pityocampa) (Battisti et al., 2005; Assefa et al., 2015). These species can maintain activity during warm conditions and therefore have relatively longer flight periods, which can elevate dispersal rates (Zera & Denno, 1997). Population expansion appears to be more likely for winged ectotherms such as lepidopterans (Cudmore et al., 2010; Mason et al., 2019), compared to apterous endothermic species (Zhu, Woodall & Clark, 2012). In some instances, lepidopteran populations are performing better than expected (Hunter et al., 2014), especially in complex ecological systems such as those in the tropics (Cheng et al., 2019).
Diet specialization is a costly life‐history strategy under a changing climate, since specialists are less likely to find suitable habitat patches during range shifts. There are a few cases of range expansion in specialists when their diets consist of high‐nitrogen resources (Betzholtz et al., 2012). In most cases, oligo‐ or polyphagous species (diet generalists) have an advantage because they have a wider breadth of host plants, allowing for easy colonization of new sites during range expansion (Betzholtz et al., 2012). By contrast, diet and habitat specialists typically have poor dispersal ability and may not be able to track environmental changes when suitable habitat patches are reduced and fragmented, resulting in local extinctions and range declines (Warren et al., 2001).
(2). Population and range retraction
While poleward expansions due to climate change are well documented in the literature, there is also evidence for population declines and local extinctions, especially for species with ranges that are limited by elevation. A survey of four butterfly species at 421 sites in the northern mountains of the UK showed that future climate warming jeopardizes the survival of all the species examined (Franco et al., 2006). Montane species are especially vulnerable to climate change impacts because mountains are predicted to be subjected to extreme temperature increases (Nogués‐Bravo et al., 2007; Molina‐Martínez et al., 2016) and because altitudinal shifts of low‐elevation species at range boundaries may confine them to more fragmented habitats (Wilson et al., 2005). However, if cooler microclimates are available at higher elevations, cold‐adapted species could remain in their optimal thermal window by shifting their range altitudinally rather than latitudinally (Beckage et al., 2008). Since elevational temperature gradients in tropical regions have a stronger effect on species' distributions than latitudinal gradients (Colwell et al., 2008), with some exceptions (Braby et al., 2014), it is likely that tropical organisms will respond to increasing temperatures with uphill shifts (Pounds, Fogden & Campbell, 1999; Colwell et al., 2008; Chen et al., 2011; Laurance et al., 2011; Molina‐Martínez et al., 2016). In a study in Oaxaca, Mexico, tropical butterfly species occurring above 1000 m showed more marked changes in their elevational distribution compared to lower altitude species (Wilson et al., 2005). Alpine ecosystems are one of the most threatened habitats under climate change. Invasive species are shifting further into these habitats and may have severe impacts on lepidopteran richness and species composition at local and regional scales (Bílá et al., 2016).
In North America, Edith's checkerspot (E. editha) provides one of the best‐documented cases of population‐level extinctions (Thomas, Franco & Hill, 2006). Extinctions of two populations of E. editha were caused by both habitat loss and regional climate change, including increasing variability in precipitation (McLaughlin et al., 2002). The West Virginia white butterfly (Pieris virginiensis) faces critical habitat loss due to climate change and is threatened by the introduction of the non‐native herb Alliaria petiolata, on which it lays two‐thirds of its eggs despite its unsuitability as a host plant (Davis et al., 2016). In the UK, fine‐resolution (1 km2) mapping demonstrated extinction gradients for three of four northern butterfly species, a lycaenid (Aricia artaxerxes) and two nymphalid species, Erebia aethiops and E. epiphron (Franco et al., 2006). Although several studies have predicted and observed range retractions under climate change, further research is needed to assess impacts on individual populations since different species are likely to respond in individual ways to climate change, and new species associations will develop as climate warming continues (Huntley et al., 1995; Hill, Thomas & Huntley, 1999b ).
IV. PHENOLOGICAL RESPONSES
(1). Voltinism
With climate change altering the timing and characteristics of seasons worldwide, adaptation will be required for many species that express different traits during different times of the year, e.g. wet‐ and dry‐season forms of multigenerational butterflies. Numerous butterfly and moth species have shifted their seasonal activities in response to variation in climatic patterns due to climate change (Pelini et al., 2009). These shifts are linked to environmental suitability since many species generally have a restricted time period in the year in which to reproduce under favourable conditions (Chuang & Peterson, 2016). The number of generations per year (voltinism) can be critical for survival of a species because an additional generation per unit time may accelerate population growth (Altermatt, 2010a ). Many species have more than one brood per year (multivoltine), and can express two developmental pathways: accelerated growth or diapause that delays development until environmental conditions are suitable (typical in temperate insects to survive the winter) (Altermatt, 2010a ). Multivoltine species are more likely to be successful under warmer temperatures. In northern European moth communities, increased multivoltinism has been observed in response to increased temperatures in the spring and summer months (Pöyry et al., 2011). The benefit of multivoltinism may be even greater when phenological changes occur earlier. In a study of 130 British moths and butterflies, multivoltine species exhibited population increases and range expansions over 19 years, while univoltine species exhibited the opposite trends (Macgregor et al., 2019a ).
In many cases, climate change‐mediated shifts in voltinism are associated with species' life cycles and larval diet spectrum (Altermatt, 2010b ). For example, a study spanning 150 years on 566 European moth and butterfly species showed that, in response to climate change, species that fed on herbaceous plants demonstrated increased voltinism but smaller shifts in flight periods compared to species feeding on woody plants (Altermatt, 2010b ). Multivoltinism has also been linked to pest outbreaks and could have consequences on agriculture and forestry (Altermatt, 2010a ). The codling moth, Cydia pomonella, is a global pest in apple fruit production. In response to higher temperatures early in the year, it is able to increase its voltinism by adding an additional generation (El Iraqui & Hmimina, 2016).
When responding to climate change, shifts towards bi‐ or multivoltinism may not always be beneficial. Some butterflies or moths that reproduce multiple times per year may be more susceptible to climate change if the negative effects accumulate over several generations. In one study monitoring 82 butterfly species in the Mediterranean throughout the year, multivoltine species had the highest population fluctuations within seasons, and suffered population declines due to extreme drought events during late‐summer generations (Melero, Stefanescu & Pino, 2016). Rapid environmental changes may also distort environmental cues that species use to time developmental decisions before winter. Species that reproduce multiple times per year use locally adapted photoperiod cues (Lindestad et al., 2019), and have strong thermal plasticity may be particularly vulnerable to developmental traps (Van Dyck et al., 2015). Overall fitness may be greatly reduced if developmental synchrony associated with multivoltinism and host plant phenology is disrupted (Altermatt, 2010a ). Local populations and species interactions are expected to be significantly impacted by these phenological changes (Altermatt, 2010b ; Forrest, 2016).
(2). Early emergence and asynchronies
Temporal mismatches with food resources is a major concern under climate change, and species will likely vary in their responses. Within Lepidoptera, differences in the magnitude of phenological shifts are correlated with various life‐history traits, including seasonal appearance, overwintering stage (diapause), food availability, habitat, altitude, and latitude (Altermatt, 2010a , 2012; Diamond et al., 2011; Karlsson, 2014; Navarro‐Cano et al., 2015). Temperature increases alter the development of insect herbivores more than their hosts, leading to insect–plant asynchronies (Dewar & Watt, 1992; Pelini et al., 2009). However, temperature is not the sole driver of phenological mismatch – increased variability in precipitation may also lead to population declines due to the timing of plant senescence relative to larval development (McLaughlin et al., 2002). Many butterfly species have been recorded emerging earlier from diapause due to advanced springs, which may lead to larval die‐off from subsequent returns to winter conditions or insufficient resources from slower host‐plant responses (e.g. flowering phenology) (Sparks, Roy & Dennis, 2005; Parmesan, 2007). Species that overwinter in the pupal stage typically advance their phenology more than species that overwinter in the larval or egg stage (Altermatt, 2010a ; Diamond et al., 2011; Karlsson, 2014), indicating that species with overwintering pupae may be at greatest risk of climate change‐related extinctions.
Indirect effects of fluctuating climates may also lead to asynchrony. Timing of snowmelt is an important trigger for alpine and temperate species to end diapause and influences population growth. For example, in Greenland, the dynamics of snowmelt and increased temperatures were correlated with the timing of onset, and the peak and end of the flight season of the Arctic butterflies B. chariclea and C. hecla (Høye et al., 2014). Similarly, early spring snow melt from increased temperatures impacts the population dynamics of the Mormon fritillary (Speyeria mormonia) through direct effects on floral abundance impacting nectar availability, leading to decreased fecundity and offspring survival as larvae and pupae in the subsequent season (Boggs & Inouye, 2012).
Butterflies and moths are significantly affected by the availability and quality of host plants as they shift geographically due to warming temperatures (Pelini et al., 2010). Unsynchronized geographic shifts between lepidopterans and their host plants may occur during climate change since they tend to disperse at different rates (Schweiger et al., 2008; Pelini et al., 2010; Posledovich et al., 2018). Thus, unequal changes in the distribution of butterflies and moths and their host plants can cause mismatches and prevent butterfly and moth populations from establishing in new areas (Pelini et al., 2010; Altermatt, 2010b , 2012; Diamond et al., 2011; Karlsson, 2014; Navarro‐Cano et al., 2015).
Phenological plasticity has been observed in many species and is considered an indicator of resilience to climate change impacts. The species that are capable of shifting their phenology may be more successful than those that are not as adaptable (Willis et al., 2010; Cleland et al., 2012; Polgar et al., 2013). For instance, lycaenid butterflies in the northeastern USA are responding to climate change by flying earlier in warmer years (Polgar et al., 2013). The orange tip butterfly (A. cardamines) in the Netherlands has advanced its first appearance in response to earlier warmer temperatures in the spring (Van Der Kolk, Wallis DeVries & Van Vliet, 2016). This species has large latitudinal variation in host use, allowing it to mediate climate change by switching among plant species based on their flowering phenology (Van Der Kolk et al., 2016) since the larvae feed on developing seed pods (Dempster, 1997). In general, species that feed on particular phenological (i.e. developmental) stages of their host plants (flower buds, young fruits, seeds, or young leaves) – termed phenological specialists (Navarro‐Cano et al., 2015; Stålhandske et al., 2016) – may be buffered against phenological mismatches if they are also host generalists (Navarro‐Cano et al., 2015). However, phenological specialization could present challenges for host specialists since they cannot adjust for phenological mismatches by switching hosts (Navarro‐Cano et al., 2015; Davies, 2019). Further research is needed on individual lepidopteran species and their host preferences, especially along latitudinal gradients, to understand fully how species interactions are impacted by climate change (Navarro‐Cano et al., 2015). Additionally, future research should focus on recent shifts and adaptive evolution of host plants due to global warming, since relatively little is known about this (Macel et al., 2017).
V. COMMUNITY RESPONSES (SPECIES INTERACTIONS)
(1). Trophic mismatches
The impacts of climate change on multi‐trophic interactions are the most complex and likely the most important responses to understand. A study by Macgregor et al. (2019b ) found no link between changes in moth biomass and ecosystem, mean precipitation, temperature, or plant productivity, possibly indicating that biomass changes can be instead attributed to asynchronies between host plants and predators resulting from climatic variability. Decoupled interactions between butterflies, their resources, and their predators have been observed in response to climate change in several cases (Visser, Bath & Lambrechts, 2004; Stireman et al., 2005). For example, Hunter et al. (2014) found that larval moths that fed on non‐vascular plants were likely to show negative responses during projected climate changes. Another notable mismatch has been seen between an insectivorous bird, the great tit (Parus major), which feeds its offspring on winter moth (Operophtera brumata) and oak leaf roller (Tortrix viridana) caterpillars. Peak caterpillar biomass of these two species has advanced earlier in the season, so this bird species may need to breed earlier to align with peak caterpillar abundance, or find other available foods (Visser, Holleman & Gienapp, 2006). Since the populations of both these moths can cause serious pest outbreaks, their decoupling from an important regulatory predator may have serious economic consequences.
Variation in environmental and climatic conditions can also lead to rapid changes in lepidopteran–parasitoid synchrony (Singer & Parmesan, 2010). Based on a comparison of 15 databases from geographically dispersed Lepidoptera rearing programs ranging from southern Canada to central Brazil, levels of parasitism in caterpillars decreased as climatic variability increased (Singer & Parmesan, 2010). This decrease in parasitism intensifies the frequency of lepidopteran larval outbreaks since parasitoids play a strong role in regulating insect herbivore populations, and climatic variability impairs the ability of parasitoids to track hosts (Stireman et al., 2005). Escape from parasitism may be important in range shifts, as demonstrated in the brown argus butterfly (A. agestis). In newly colonized areas, this species has lower mortality from parasitoids (Menéndez et al., 2008) which has contributed to its expansion northward in the UK over the last 30 years. Climate change could also affect parasitism via changes in behaviour. Larvae of the Glanville fritillary (M. cinxia) use solar radiation to outpace a braconid wasp parasitoid, Cotesia melitaearum. Warmer temperatures create asynchrony by enabling faster developmental rates in caterpillars than their parasitoids (Van Nouhuys & Lei, 2004). By understanding the role of environmental variability on species interactions, we thus can gain insight into how the effects of climate change could impact larger processes at community and ecosystem levels (Jamieson et al., 2012).
It is evident that many species differ in their physiological tolerances to varying climatic patterns, which may lead to asynchrony between trophic levels (Parmesan, 2006; Schweiger et al., 2008; Filz & Schmitt, 2015). Species that rely on specific obligate biotic interactions, such as mutualism and parasitism, are of highest concern. Over the last 20 years, the largest declines in European butterflies have been demonstrated in myrmecophilic species (Thomas, 2005). Many species of Lycaenidae depend on ants for survival (Pierce et al., 2002) and environmental changes might result in severe asynchrony between the butterflies and their hosts. In particular, Phengaris butterflies depend on Myrmica ants for the survival of their offspring. A study that evaluated niche overlap and host specificity for Phengaris butterflies and Myrmica ants under climate change scenarios revealed that the butterflies should retain their connection to their hosts and maintain similar niche overlap over time (Filz & Schmitt, 2015). However, these butterflies are also threatened by severe habitat loss, and conservation management plans should include the host ants and their current and future niches (Filz & Schmitt, 2015), which may affect their resilience as mutualists (Rafferty, CaraDonna & Bronstein, 2015). Similar studies need to assess climate change impacts on other endangered lycaenid butterflies and their host ants.
VI. APPLYING THE LEPIDOPTERA CLIMATE CHANGE MODEL
In our review we identified general patterns in how animals may respond to climate change by using the well‐studied Lepidoptera as a model taxon. One of the most frequent conclusions within the Lepidoptera literature is that phenotypic plasticity allows species to cope with changing climate. Phenotypic plasticity in physiology, behaviour, and phenology across developmental stages, habitats, and phylogeny typically confers resilience to climate change in Lepidoptera. Similar patterns have been found in other plants and animals (Charmantier et al., 2008; Nicotra et al., 2010; Seebacher, White & Franklin, 2015), suggesting that this may be a good indicator of species resilience. However, we caution using this as a rule; there are instances where ecological specialization can be a benefit under specific circumstances (Duputié et al., 2015; Oostra et al., 2018).
It has repeatedly been shown that climate change is altering the distribution of lepidopteran populations, with population retraction and local extinction being the most common responses (Lenoir et al., 2020). This phenomenon is also repeatedly found in non‐lepidopteran taxa, including plants, invertebrates, and vertebrates spanning diverse terrestrial, freshwater, and marine ecosystems (Maclean & Wilson, 2011; Cahill et al., 2013; Selwood, McGeoch & MacNally, 2015). These broad trends within and across taxa strengthen the need for an increased effort toward climate change mitigation, and increased research on conservation strategies. It is worth noting that increased research on species ecology carries conservation benefits in the context of habitat loss and fragmentation, which are causing extinctions regardless of climate change.
The race to save imperilled species is likely to depend on the implementation of conservation genetics in the context of species‐specific ecological requirements. As discussed in Section II.1, Pgi and Hsp70 have been used in Lepidoptera to detect responses to climate change; microsatellites, functional genes, and other markers have proved to be similarly useful in many plants and animals (Vandergast et al., 2008; Aitken & Whitlock, 2013; Fordham et al., 2014; Franks et al., 2014). Further research is needed to identify markers from diverse phylogenetic groups. However, the efficacy of genetic markers will depend on whether specific ecological information is available: a thorough understanding of a species' temperature, behavioural, and habitat preferences must be considered in conjunction with predicted gene functionality in altered environments for conservation efforts under climate change. Since ecological specialists are often the most threatened, carefully targeted ecological research will be required to determine the best conservation approaches. It thus is critical to continue ecological monitoring alongside implementation of habitat conservation and climate change mitigation practices (Thomas, 2016).
The impacts of climate change on community interactions are usually the hardest to detect, yet are likely one of the most important topics to address. It is evident from research across butterflies and moths that climate change can lead to detrimental impacts associated with symbiont and resource asynchronies. A similar impact is also foreseen in other plant and animal communities, with cascading ecological and evolutionary effects that become increasingly difficult to predict (Winder & Schindler, 2004; Lavergne et al., 2010; Warren & Bradford, 2014; Classen et al., 2015; Chidawanyika, Mudavanhu & Nyamukondiwa, 2019). Many of these studies identify ecological specialization and symbiont fidelity as risk factors for adapting to climate change. Future research should integrate symbionts and their interactions within communities into conservation strategies.
VII. CONCLUSIONS
We examined the impacts of climate change on butterflies and moths. The effects of climate change on lepidopterans include: selection on dispersal (e.g. Pgi) genotypes, up‐regulation of thermoregulation (e.g. heat shock protein) genes, decreased melanism, reduced body size, increased dispersal, shifts between microhabitats and across altitude and latitude, habitat and host generalization, a shift towards multiple generations per year, and early emergence from diapause.
The impact of climate change on the genetics, physiology, behaviour, and morphology of insects is understudied. The limited research that has been conducted focuses on a few species. Future research should be expanded to include additional taxa that are ecologically and phylogenetically divergent, to understand the effects of climate change.
Many species are expanding polewards to escape warming temperatures. Thermophilization and diet specialization may impact this process by affecting their ability to invade new habitats. Future research should focus on individual populations since these are likely to show local adaptation and hence unique responses to climate change.
There is much research available on phenology in a climate change framework for Lepidoptera. The majority of studies report early emergence from diapause and increasing asynchrony with host plants. This may be particularly crucial for specialists that use only a particular part of a plant, or a particular stage in its life history, and are therefore phenologically constrained – such as lycaenid butterflies that specialize on flower buds or seeds. Other climatic changes such as early snow melt and shifts in precipitation patterns have led to lepidopteran population declines and local extinctions. Changes to voltinism are common, with numerous species increasing the number of generations per year. Future research should investigate how host preferences are likely to change in relation to predicted phenological shifts due to climate change.
VIII. ACKNOWLEDGEMENTS
We thank U. Somjee and anonymous reviewers for comments and discussion, and X. Zheng for assistance with the figure. We acknowledge Graduate Research Assistantship support (to G.M.H.) from the Disney Conservation Fund. The authors declare no conflict of interest.
REFERENCES
- Aalberg Haugen, I. M. & Gotthard, K. (2015). Diapause induction and relaxed selection on alternative developmental pathways in a butterfly. Journal of Animal Ecology 84(2), 464–472. [DOI] [PubMed] [Google Scholar]
- Aitken, S. N. & Whitlock, M. C. (2013). Assisted gene flow to facilitate local adaptation to climate change. Annual Review of Ecology, Evolution, and Systematics 44, 367–388. [Google Scholar]
- Altermatt, F. (2010a). Climatic warming increases voltinism in European butterflies and moths. Proceedings of the Royal Society of London Series B: Biological Sciences 277, 1281–1287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altermatt, F. (2010b). Tell me what you eat and I'll tell you when you fly: diet can predict phenological changes in response to climate change. Ecology Letters 13(12), 1475–1484. [DOI] [PubMed] [Google Scholar]
- Altermatt, F. (2012). Temperature‐related shifts in butterfly phenology depend on the habitat. Global Change Biology 18(8), 2429–2438. [Google Scholar]
- Ashton, S. , Gutiérrez, D. & Wilson, R. J. (2009). Effects of temperature and elevation on habitat use by a rare mountain butterfly: implications for species responses to climate change. Ecological Entomology 34(4), 437–446. [Google Scholar]
- Assefa, Y. , Conlong, D. E. , Van Den Berg, J. & Martin, L. A. (2015). Ecological genetics and host range expansion by Busseola fusca (Lepidoptera: Noctuidae). Environmental Entomology 44(4), 1265–1274. [DOI] [PubMed] [Google Scholar]
- Bahar, M. H. , Hegedus, D. , Soroka, J. , Coutu, C. , Bekkaoui, D. & Dosdall, L. (2013). Survival and Hsp70 gene expression in Plutella xylostella and its larval parasitoid Diadegma insulare varied between slowly ramping and abrupt extreme temperature regimes. PLoS One 8(9), 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bar‐On, Y. M. , Phillips, R. & Milo, R. (2018). The biomass distribution on Earth. Proceedings of the National Academy of Sciences of the United States of America 115(25), 6506–6511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Battisti, A. , Stastny, M. , Netherer, S. , Robinet, C. , Schopf, A. , Roques, A. & Larsson, S. (2005). Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecological Applications 15(6), 2084–2096. [Google Scholar]
- Beckage, B. , Osborne, B. , Gavin, D. G. , Pucko, C. , Siccama, T. & Perkins, T. (2008). A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proceedings of the National Academy of Sciences of the United States of America 105(11), 4197–4202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belitz, M. W. , Hendrick, L. K. , Monfils, M. J. , Cuthrell, D. L. , Marshall, C. J. , Kawahara, A. Y. , Cobb, N. S. , Zaspel, J. M. , Horton, A. M. , Huber, S. L. & Warren, A. D. (2018). Aggregated occurrence records of the federally endangered Poweshiek skipperling (Oarisma poweshiek). Biodiversity Data Journal 6, e29081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Betzholtz, P. , Pettersson, L. B. , Ryrholm, N. & Franze, M. (2012). With that diet, you will go far: trait‐based analysis reveals a link between rapid range expansion and a nitrogen‐favoured diet. Proceedings of the Royal Society B: Biological Sciences 280, 20122305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bílá, K. , Šipoš, J. , Kindlmann, P. & Kuras, T. (2016). Consequences for selected high‐elevation butterflies and moths from the spread of Pinus mugo into the alpine zone in the High Sudetes Mountains. PeerJ 4, e2094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boggs, C. L. & Inouye, D. W. (2012). A single climate driver has direct and indirect effects on insect population dynamics. Ecology Letters 15(5), 502–508. [DOI] [PubMed] [Google Scholar]
- Bonebrake, T. C. , Boggs, C. L. , Stamberger, J. A. , Deutsch, C. A. & Ehrlich, P. R. (2014). From global change to a butterfly flapping: biophysics and behaviour affect tropical climate change impacts. Proceedings of the Royal Society of London B: Biological Sciences 281(1793), 20141264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowden, J. J. , Eskildsen, A. , Hansen, R. R. , Olsen, K. , Kurle, C. M. & Høye, T. T. (2015). High‐Arctic butterflies become smaller with rising temperatures. Biology Letters 11(10), 1–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braby, M. F. , Bertelsmeier, C. , Sanderson, C. & Thistleton, B. M. (2014). Spatial distribution and range expansion of the Tawny Coster butterfly, Acraea terpsicore (Linnaeus, 1758) (Lepidoptera: Nymphalidae), in South‐East Asia and Australia. Insect Conservation and Diversity 7(2), 132–143. [Google Scholar]
- Breed, G. A. , Stichter, S. & Crone, E. E. (2013). Climate‐driven changes in northeastern US butterfly communities. Nature Climate Change 3(2), 142–145. [Google Scholar]
- Bryant, S. R. , Thomas, C. D. & Bale, J. S. (2000). Thermal ecology of gregarious and solitary nettle‐feeding nymphalid butterfly larvae. Oecologia 122(1), 1–10. [DOI] [PubMed] [Google Scholar]
- Cahill, A. E. , Aiello‐Lammens, M. E. , Fisher‐Reid, M. C. , Hua, X. , Karanewsky, C. J. , Yeong Ryu, H. , Sbeglia, G. C. , Spagnolo, F. , Waldron, J. B. , Warsi, O. & Wiens, J. J. (2013). How does climate change cause extinction? Proceedings of the Royal Society B: Biological Sciences 280(1750), 20121890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canale, C. I. & Henry, P. Y. (2010). Adaptive phenotypic plasticity and resilience of vertebrates to increasing climatic unpredictability. Climate Research 43(1–2), 135–147. [Google Scholar]
- Charmantier, A. , McCleery, R. H. , Cole, L. R. , Perrins, C. , Kruuk, L. E. & Sheldon, B. C. (2008). Adaptive phenotypic plasticity in response to climate change in a wild bird population. Science 320(5877), 800–803. [DOI] [PubMed] [Google Scholar]
- Chen, I.‐C. , Hill, J. K. , Shiu, H.‐J. , Holloway, J. D. , Benedick, S. , Chey, V. K. , Barlow, H. S. & Thomas, C. D. (2011). Asymmetric boundary shifts of tropical montane Lepidoptera over four decades of climate warming. Global Ecology and Biogeography 20(1), 34–45. [Google Scholar]
- Cheng, W. , Kendrick, R. C. , Guo, F. , Xing, S. , Tingley, M. W. & Bonebrake, T. C. (2019). Complex elevational shifts in a tropical lowland moth community following a decade of climate change. Diversity and Distributions 25(4), 514–523. [Google Scholar]
- Chidawanyika, F. , Mudavanhu, P. & Nyamukondiwa, C. (2019). Global climate change as a driver of bottom‐up and top‐down factors in agricultural landscapes and the fate of host‐parasitoid interactions. Frontiers in Ecology and Evolution 7, 1–13. [Google Scholar]
- Chuang, A. & Peterson, C. R. (2016). Expanding population edges: theories, traits, and trade‐offs. Global Change Biology 22(2), 494–512. [DOI] [PubMed] [Google Scholar]
- Classen, A. T. , Sundqvist, M. K. , Henning, J. A. , Newman, G. S. , Moore, J. A. , Cregger, M. A. , Moorhead, L. C. & Patterson, C. M. (2015). Direct and indirect effects of climate change on soil microbial and soil microbial‐plant interactions: what lies ahead? Ecosphere 6(8), 1–21. [Google Scholar]
- Cleland, E. E. , Allen, J. M. , Crimmins, T. M. , Dunne, J. A. , Pau, S. , Travers, S. E. , Zavaleta, E. S. & Wolkovich, E. M. (2012). Phenological tracking enables positive species responses to climate change. Ecology 93(8), 1765–1771. [DOI] [PubMed] [Google Scholar]
- Clusella‐Trullas, S. & Nielsen, M. (2020). The evolution of insect body coloration under changing climates. Current Opinion in Insect Science 41, 25–32. [DOI] [PubMed] [Google Scholar]
- Cohen, J. M. , Lajeunesse, M. J. & Rohr, J. R. (2018). A global synthesis of animal phenological responses to climate change. Nature Climate Change 8(3), 224–228. [Google Scholar]
- Colwell, R. K. , Brehm, G. , Cardelús, C. L. , Gilman, A. C. & Longino, J. T. (2008). Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science 322(5899), 258–261. [DOI] [PubMed] [Google Scholar]
- Coulthard, E. , Norrey, J. , Shortall, C. & Harris, W. E. (2019). Ecological traits predict population changes in moths. Biological Conservation 233, 213–219. [Google Scholar]
- Crozier, L. (2003). Winter warming facilitates range expansion: cold tolerance of the butterfly Atalopedes campestris . Oecologia 135(4), 648–656. [DOI] [PubMed] [Google Scholar]
- Cudmore, T. J. , Björklund, N. , Carroll, A. L. & Lindgren, B. S. (2010). Climate change and range expansion of an aggressive bark beetle: evidence of higher beetle reproduction in naïve host tree populations. Journal of Applied Ecology 47(5), 1036–1043. [Google Scholar]
- Davies, W. J. (2019). Multiple temperature effects on phenology and body size in wild butterflies predict a complex response to climate change. Ecology 100(4), e02612. [DOI] [PubMed] [Google Scholar]
- Davies, W. J. & Saccheri, I. J. (2013). Maintenance of body‐size variation and host range in the orange‐tip butterfly: evidence for a trade‐off between adult life‐history traits. Ecological Entomology 38, 49–60. [Google Scholar]
- Davis, S. L. , Cipollini, D. , Didham, R. & Littlewood, N. (2016). Range, genetic diversity and future of the threatened butterfly, Pieris virginiensis . Insect Conservation and Diversity 9(6), 506–516. [Google Scholar]
- Dempster, J. P. (1997). The role of larval food resources and adult movement in the population dynamics of the orange‐tip butterfly (Anthocharis cardamines). Oecologia 111(4), 549–556. [DOI] [PubMed] [Google Scholar]
- Dempster, J. P. , King, M. L. & Lakhani, K. H. (1976). The status of the swallowtail butterfly in Britain. Ecological Entomology 1(2), 71–84. [Google Scholar]
- Dewar, R. C. & Watt, A. D. (1992). Predicted changes in the synchrony of larval emergence and budburst under climatic warming. Oecologia 9, 557–559. [DOI] [PubMed] [Google Scholar]
- Diamond, S. E. , Frame, A. M. , Martin, R. A. & Buckley, L. B. (2011). Species' traits predict phenological responses to climate change in butterflies. Ecology 92(5), 1005–1012. [DOI] [PubMed] [Google Scholar]
- Dukes, J. S. , Pontius, J. , Orwig, D. , Garnas, J. R. , Rodgers, V. L. , Brazee, N. , Cooke, B. , Theoharides, K. A. , Stange, E. E. , Harrington, R. , Ehrenfeld, J. , Gurevitch, J. , Lerdau, M. , Stinson, K. , Wick, R. , et al. (2009). Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America. Canadian Journal of Forest Research 39(2), 231–248. [Google Scholar]
- Duputié, A. , Rutschmann, A. , Ronce, O. & Chuine, I. (2015). Phenological plasticity will not help all species adapt to climate change. Global Change Biology 21(8), 3062–3073. [DOI] [PubMed] [Google Scholar]
- Duque, A. , Stevenson, P. R. & Feeley, K. J. (2015). Thermophilization of adult and juvenile tree communities in the northern tropical Andes. Proceedings of the National Academy of Sciences 112(34), 10744–10749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El Iraqui, S. & Hmimina, M. (2016). Impact of temperatures on the voltinism of Cydia pomonella (Lepidoptera: Tortricidae). Annals of the Entomological Society of America 109(5), 698–704. [Google Scholar]
- Ellis, S. , Wainwright, D. , Dennis, E. B. , Bourn, N. A. D. , Bulman, C. R. , Hobson, R. , Jones, R. , Middlebrook, I. , Plackett, J. , Smith, R. G. & Wain, M. (2019). Are habitat changes driving the decline of the UK's most threatened butterfly: the High Brown Fritillary Argynnis adippe (Lepidoptera: Nymphalidae)? Journal of Insect Conservation 23(2), 351–367. [Google Scholar]
- Fenberg, P. B. , Self, A. , Stewart, J. R. , Wilson, R. J. & Brooks, S. J. (2016). Exploring the universal ecological responses to climate change in a univoltine butterfly. Journal of Animal Ecology 85(3), 739–748. [DOI] [PubMed] [Google Scholar]
- Filz, K. J. & Schmitt, T. (2015). Niche overlap and host specificity in parasitic Maculinea butterflies (Lepidoptera: Lycaenidae) as a measure for potential extinction risks under climate change. Organisms Diversity & Evolution 15(3), 555–565. [Google Scholar]
- Finkbeiner, S. D. , Reed, R. D. , Dirig, R. & Losey, J. E. (2011). The role of environmental factors in the northeastern range expansion of Papilio cresphontes . Journal of the Lepidopterists' Society 65(2), 119–125. [Google Scholar]
- Fischer, K. & Karl, I. (2010). Exploring Plastic and genetic responses to temperature variation using copper butterflies. Climate Research 43, 17–30. [Google Scholar]
- Fordham, D. A. , Brook, B. W. , Moritz, C. & Nogués‐Bravo, D. (2014). Better forecasts of range dynamics using genetic data. Trends in Ecology & Evolution 29(8), 436–443. [DOI] [PubMed] [Google Scholar]
- Forister, M. L. , McCall, A. C. , Sanders, N. J. , Fordyce, J. A. , Thorne, J. H. , O'Brien, J. , Waetjen, D. P. & Shapiro, A. M. (2010). Compounded effects of climate change and habitat alteration shift patterns of butterfly diversity. Proceedings of the National Academy of Sciences 107(5), 2088–2092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forister, M. L. , Pelton, E. M. & Black, S. H. (2019). Declines in insect abundance and diversity: we know enough to act now. Conservation Science and Practice 1(8), e80. [Google Scholar]
- Forrest, J. R. (2016). Complex responses of insect phenology to climate change. Current Opinion in Insect Science 17, 49–54. [DOI] [PubMed] [Google Scholar]
- Forsman, A. , Betzholtz, P.‐E. & Franzén, M. (2016). Faster poleward range shifts in moths with more variable colour patterns. Scientific Reports 6, 36265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox, R. (2013). The decline of moths in Great Britain: a review of possible causes. Insect Conservation and Diversity 6, 5–19. [Google Scholar]
- Franco, A. M. A. , Hill, J. K. , Kitschke, C. , Collingham, Y. C. , Roy, D. B. , Fox, R. , Huntley, B. & Thomas, C. D. (2006). Impacts of climate warming and habitat loss on extinctions at species' low‐latitude range boundaries. Global Change Biology 12(8), 1545–1553. [Google Scholar]
- Franks, S. J. , Weber, J. J. & Aitken, S. N. (2014). Evolutionary and plastic responses to climate change in terrestrial plant populations. Evolutionary Applications 7(1), 123–139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottfried, M. , Pauli, H. , Futschik, A. , Akhalkatsi, M. , Barančok, P. , Benito Alonso, J. L. , Coldeo, G. , Dick, J. , Erschbamer, B. , Calzado, M. R. F. , Kazakis, G. , Krajči, J. , Larsson, P. , Mallaun, M. , Michelsen, O. , et al. (2012). Continent‐wide response of mountain vegetation to climate change. Nature Climate Change 2(2), 111–115. [Google Scholar]
- Haag, C. R. , Saastamoinen, M. , Marden, J. H. & Hanski, I. (2005). A candidate locus for variation in dispersal rate in a butterfly metapopulation. Proceedings of the Royal Society B: Biological Sciences 272(1580), 2449–2456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halsch, C. A. , Shapiro, A. M. , Fordyce, J. A. , Nice, C. C. , Thorne, J. H. , Waetjen, D. P. & Forister, M. L. (2021). Insects and recent climate change. Proceedings of the National Academy of Sciences of the United States of America 118(2), e2002543117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanski, I. , Breuker, C. J. , Schops, K. , Setchfield, R. & Nieminen, M. (2002). Population history and life history influence the migration rate of female Glanville fritillary butterflies. Oikos 98(1), 87–97. [Google Scholar]
- Hanski, I. , Saastamoinen, M. & Ovaskainen, O. (2006). Dispersal‐related life‐history trade‐offs in a butterfly metapopulation. Journal of Animal Ecology 75(1), 91–100. [DOI] [PubMed] [Google Scholar]
- Hanski, I. & Saccheri, I. (2006). Molecular‐level variation affects population growth in a butterfly metapopulation. PLoS Biology 4(5), 719–726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawksworth, D. L. & Lücking, R. (2017). Fungal diversity revisited: 2.2 to 3.8 million species. In Heitman, J , Howlett, B. J. , Crous, P. W. , Stukenbrock, E. H. , James, T. Y. & Gow, N. A. R. , The Fungal Kingdom. Washington D.C: American Society for Microbiology, pp. 79–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hellmann, J. J. , Pelini, S. L. , Prior, K. M. & Dzurisin, J. D. K. (2008). The response of two butterfly species to climatic variation at the edge of their range and the implications for poleward range shifts. Oecologia 157(4), 583–592. [DOI] [PubMed] [Google Scholar]
- Hill, A. J. K. , Thomas, C. D. & Blakeley, D. S. (1999a). Evolution of flight morphology in a butterfly that has recently expanded its geographic range. Oecologia 121, 165–170. [DOI] [PubMed] [Google Scholar]
- Hill, J. K. , Thomas, C. D. & Huntley, B. (1999b). Climate and habitat availability determine 20th century change in a butterfly's range margin. Proceedings of the Royal Society of London. Series B: Biological Sciences 266, 1197–1206. [Google Scholar]
- Hoffmann, A. A. & Daborn, P. J. (2007). Towards genetic markers in animal populations as biomonitors for human‐induced environmental change. Ecology Letters 10(1), 63–76. [DOI] [PubMed] [Google Scholar]
- Høye, T. T. , Eskildsen, A. , Hansen, R. R. , Bowden, J. J. , Schmidt, N. M. & Kissling, W. D. (2014). Phenology of high‐arctic butterflies and their floral resources: species‐specific responses to climate change. Current Zoology 60(2), 243–251. [Google Scholar]
- Hughes, C. L. , Dytham, C. & Hill, J. K. (2007). Modelling and analysing evolution of dispersal in populations at expanding range boundaries. Ecological Entomology 32(5), 437–445. [Google Scholar]
- Hunter, M. D. , Kozlov, M. V. , Itämies, J. , Pulliainen, E. , Bäck, J. , Kyrö, E. M. & Niemelä, P. (2014). Current temporal trends in moth abundance are counter to predicted effects of climate change in an assemblage of subarctic forest moths. Global Change Biology 20(6), 1723–1737. [DOI] [PubMed] [Google Scholar]
- Huntley, B. , Berry, P. M. , Cramer, W. & Mcdonald, A. P. (1995). Modelling present and potential future ranges of some European higher plants using climate response surfaces. Journal of Biogeography 22(6), 967–1001. [Google Scholar]
- Jamieson, M. A. , Trowbridge, A. M. , Raffa, K. F. & Lindroth, R. L. (2012). Consequences of climate warming and altered precipitation patterns for plant‐insect and multitrophic interactions. Plant Physiology 160(4), 1719–1727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kallioniemi, E. & Hanski, I. (2011). Interactive effects of Pgi genotype and temperature on larval growth and survival in the Glanville fritillary butterfly. Functional Ecology 25(5), 1032–1039. [Google Scholar]
- Karl, I. , Schmitt, T. & Fischer, K. (2009). Genetic differentiation between alpine and lowland populations of a butterfly is related to PGI enzyme genotype. Ecography 32(3), 488–496. [Google Scholar]
- Karlsson, B. (2014). Extended season for northern butterflies. International Journal of Biometeorology 58(5), 691–701. [DOI] [PubMed] [Google Scholar]
- Kearney, M. , Shine, R. & Porter, W. P. (2009). The potential for behavioral thermoregulation to buffer “cold‐blooded” animals against climate warming. Proceedings of the National Academy of Sciences of the United States of America 106(10), 3835–3840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kingsolver, J. G. (1985). Thermoregulatory significance of wing melanization in Pieris butterflies (Lepidoptera: Pieridae): physics, posture, and pattern. Oecologia 66(4), 546–553. [DOI] [PubMed] [Google Scholar]
- Kingsolver, J. G. & Buckley, L. B. (2014). Climate variability slows evolutionary responses of Colias butterflies to recent climate change. Proceedings of the Royal Society B: Biological Sciences 282(18020), 20142470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kingsolver, J. G. & Moffat, R. J. (1982). Thermoregulation and the determinants of heat transfer in Colias butterflies. Oecologia 53(1), 27–33. [DOI] [PubMed] [Google Scholar]
- Kiritani, K. (2006). Predicting impacts of global warming on population dynamics and distribution of arthropods in Japan. Population Ecology 48(1), 5–12. [Google Scholar]
- Kleckova, I. & Klecka, J. (2016). Facing the heat: thermoregulation and behaviour of lowland species of a cold‐dwelling butterfly genus, Erebia . PLoS One 11(3), 1–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuussaari, M. , Nieminen, M. & Hanski, I. (1996). An experimental study of migration in the Glanville fritillary butterfly Melitaea cinxia . Journal of Animal Ecology 65, 791–801. [Google Scholar]
- Laurance, W. F. , Carolina Useche, D. , Shoo, L. P. , Herzog, S. K. , Kessler, M. , Escobar, F. , Brehm, G. , Axmacher, J. C. , Ching Chen, I. , Gamez Arellano, L. , Hietz, P. , Fiedler, K. , Pyrcz, T. , Wolf, J. , Merkord, C. L. , et al. (2011). Global warming, elevational ranges and the vulnerability of tropical biota. Biological Conservation 144(1), 548–557. [Google Scholar]
- Lavergne, S. , Mouquet, N. , Thuiller, W. & Ronce, O. (2010). Biodiversity and climate change: integrating evolutionary and ecological responses of species and communities. Annual Review of Ecology, Evolution, and Systematics 41, 321–350. [Google Scholar]
- Lenoir, J. , Bertrand, R. , Comte, L. , Bourgeaud, L. , Hattab, T. , Murienne, J. & Grenouillet, G. (2020). Species better track climate warming in the oceans than on land. Nature Ecology & Evolution 4(8), 1044–1059. [DOI] [PubMed] [Google Scholar]
- Lindestad, O. , Wheat, C. W. , Nylin, S. & Gotthard, K. (2019). Local adaptation of photoperiodic plasticity maintains life cycle variation within latitudes in a butterfly. Ecology 100(1), e02550. [DOI] [PubMed] [Google Scholar]
- Lister, B. C. & Garcia, A. (2018). Climate‐driven declines in arthropod abundance restructure a rainforest food web. Proceedings of the National Academy of Sciences of the United States of America 115(44), E10397–E10406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macel, M. , Dostálek, T. , Esch, S. , Bucharová, A. , van Dam, N. M. , Tielbörger, K. , Verhoeven, K. J. F. & Münzbergová, Z. (2017). Evolutionary responses to climate change in a range expanding plant. Oecologia 184(2), 543–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macgregor, C. J. , Thomas, C. D. , Roy, D. B. , Beaumont, M. A. , Bell, J. R. , Brereton, T. , Bridle, J. R. , Dytham, C. , Fox, R. , Gotthard, K. & Hoffmann, A. A. (2019a). Climate‐induced phenology shifts linked to range expansions in species with multiple reproductive cycles per year. Nature Communications 10(1), 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macgregor, C. J. , Williams, J. H. , Bell, J. R. & Thomas, C. D. (2019b). Moth biomass increases and decreases over 50 years in Britain. Nature Ecology & Evolution 3, 1645–1649. [DOI] [PubMed] [Google Scholar]
- MacLean, H. J. , Kingsolver, J. G. & Buckley, L. B. (2016). Historical changes in thermoregulatory traits of alpine butterflies reveal complex ecological and evolutionary responses to recent climate change. Climate Change Responses 3(1), 13. [Google Scholar]
- MacLean, H. J. , Nielsen, M. E. , Kingsolver, J. G. & Buckley, L. B. (2018). Using museum specimens to track morphological shifts through climate change. Philosophical Transactions of the Royal Society B 374(1763), 20170404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maclean, I. M. & Wilson, R. J. (2011). Recent ecological responses to climate change support predictions of high extinction risk. Proceedings of the National Academy of Sciences of the United States of America 108(30), 12337–12342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason, L. R. , Green, R. E. , Howard, C. , Stephens, P. A. , Willis, S. G. , Aunins, A. , Brotons, L. , Chodkiewicz, T. , Chylarecki, P. , Escandell, V. & Foppen, R. P. (2019). Population responses of bird populations to climate change on two continents vary with species' ecological traits but not with direction of change in climate suitability. Climatic Change 157(3–4), 337–354. [Google Scholar]
- Mattila, A. L. K. (2015). Thermal biology of flight in a butterfly: genotype, flight metabolism, and environmental conditions. Ecology and Evolution 5(23), 5539–5551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maurer, J. A. , Shepard, J. H. , Crabo, L. G. , Hammond, P. C. , Zack, R. S. & Peterson, M. A. (2018). Phenological responses of 215 moth species to interannual climate variation in the Pacific Northwest from 1895 through 2013. PLoS One 13(9), e0202850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLaughlin, J. F. , Hellmann, J. J. , Boggs, C. L. & Ehrlich, P. R. (2002). Climate change hastens population extinctions. Proceedings of the National Academy of Sciences of the United States of America 99(9), 6070–6074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melero, Y. , Stefanescu, C. & Pino, J. (2016). General declines in Mediterranean butterflies over the last two decades are modulated by species traits. Biological Conservation 201, 336–342. [Google Scholar]
- Menéndez, R. , González‐Megías, A. , Lewis, O. T. , Shaw, M. R. & Thomas, C. D. (2008). Escape from natural enemies during climate‐driven range expansion: a case study. Ecological Entomology 33(3), 413–421. [Google Scholar]
- Merilä, J. & Hendry, A. P. (2014). Climate change, adaptation, and phenotypic plasticity: the problem and the evidence. Evolutionary Applications 7(1), 1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misof, B. , Liu, S. , Meusemann, K. , Peters, R. S. , Donath, A. , Mayer, C. , Frandsen, P. B. , Ware, J. , Flouri, T. , Beutel, R. G. & Niehuis, O. (2014). Phylogenomics resolves the timing and pattern of insect evolution. Science 346(6210), 763–767. [DOI] [PubMed] [Google Scholar]
- Mitikka, V. & Hanski, I. (2010). Pgi genotype influences flight metabolism at the expanding range margin of the European map butterfly. Annales Zoologici Fennici 47(1), 1–14. [Google Scholar]
- Molina‐Martínez, A. , León‐Cortés, J. L. , Regan, H. M. , Lewis, O. T. , Navarrete, D. , Caballero, U. & Luis‐Martínez, A. (2016). Changes in butterfly distributions and species assemblages on a Neotropical mountain range in response to global warming and anthropogenic land use. Diversity and Distributions 22(11), 1085–1098. [Google Scholar]
- Mora, C. , Tittensor, D. P. , Adl, S. , Simpson, A. G. & Worm, B. (2011). How many species are there on Earth and in the ocean? PLoS Biology 9(8), e1001127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munro, J. T. , Medina, I. , Walker, K. , Moussalli, A. , Kearney, M. R. , Dyer, A. G. , Garcia, J. , Rankin, K. J. & Stuart‐Fox, D. (2019). Climate is a strong predictor of near‐infrared reflectance but a poor predictor of colour in butterflies. Proceedings of the Royal Society B: Biological Sciences 286(1898), 20190234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadeau, C. P. , Urban, M. C. & Bridle, J. R. (2017). Climates past, present, and yet‐to‐come shape climate change vulnerabilities. Trends in Ecology & Evolution 32(10), 786–800. [DOI] [PubMed] [Google Scholar]
- Navarro‐Cano, J. A. , Karlsson, B. , Posledovich, D. , Toftegaard, T. , Wiklund, C. , Ehrlén, J. & Gotthard, K. (2015). Climate change, phenology, and butterfly host plant utilization. Ambio 44(1), 78–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nève, G. & Després, L. (2020). Cold adaptation across the elevation gradient in an alpine butterfly species complex. Ecological Entomology 45(5), 997–1003. [Google Scholar]
- Nice, C. C. & Fordyce, J. A. (2006). How caterpillars avoid overheating: behavioral and phenotypic plasticity of pipevine swallowtail larvae. Oecologia 146(4), 541–548. [DOI] [PubMed] [Google Scholar]
- Nicotra, A. B. , Atkin, O. K. , Bonser, S. P. , Davidson, A. M. , Finnegan, E. J. , Mathesius, U. , Poot, P. , Purugganan, M. D. , Richards, C. L. , Valladares, F. & van Kleunen, M. (2010). Plant phenotypic plasticity in a changing climate. Trends in Plant Science 15(12), 684–692. [DOI] [PubMed] [Google Scholar]
- Niitepõld, K. , Smith, A. D. , Osborne, J. L. , Reynolds, D. R. , Carreck, N. L. , Martin, A. P. , Marden, J. H. , Ovaskainen, O. & Hanski, I. (2009). Flight metabolic rate and Pgi genotype influence butterfly dispersal rate in the field. Ecology 90(8), 2223–2232. [DOI] [PubMed] [Google Scholar]
- Nogués‐Bravo, D. , Araújo, M. B. , Errea, M. P. & Martínez‐Rica, J. P. (2007). Exposure of global mountain systems to climate warming during the 21st Century. Global Environmental Change 17(3), 420–428. [Google Scholar]
- Ohlberger, J. (2013). Climate warming and ectotherm body size–from individual physiology to community ecology. Functional Ecology 27(4), 991–1001. [Google Scholar]
- Oliver, T. H. , Marshall, H. H. , Morecroft, M. D. , Brereton, T. , Prudhomme, C. & Huntingford, C. (2015). Interacting effects of climate change and habitat fragmentation on drought‐sensitive butterflies. Nature Climate Change 5(10), 941–945. [Google Scholar]
- Oostra, V. , Saastamoinen, M. , Zwaan, B. J. & Wheat, C. W. (2018). Strong phenotypic plasticity limits potential for evolutionary responses to climate change. Nature Communications 9(1), 1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parmesan, C. (2006). Ecological and evolutionary responses to recent climate change. Annual Review of Ecology, Evolution, and Systematics 37(1), 637–669. [Google Scholar]
- Parmesan, C. (2007). Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Global Change Biology 13(9), 1860–1872. [Google Scholar]
- Parmesan, C. , Ryrholm, N. , Stefanescu, C. , Hill, J. K. , Thomas, C. D. , Descimon, H. , Huntley, B. , Kaila, L. , Kullberg, J. , Tammaru, T. , Tennent, W. J. , Thomas, J. A. & Warren, M. (1999). Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 399, 579–583. [Google Scholar]
- Parmesan, C. & Yohe, G. (2003). A globally coherent fingerprint of climate change impacts across natural systems. Nature 421(6918), 37. [DOI] [PubMed] [Google Scholar]
- Pelini, S. L. , Keppel, J. A. , Kelley, A. E. & Hellmann, J. J. (2010). Adaptation to host plants may prevent rapid insect responses to climate change. Global Change Biology 16(11), 2923–2929. [Google Scholar]
- Pelini, S. L. , Prior, K. M. , Parker, D. J. , Dzurisin, J. D. K. , Lindroth, R. L. & Hellmann, J. J. (2009). Climate change and temporal and spatial mismatches in insect communities. Climate Change, 215–231. [Google Scholar]
- Pierce, N. E. , Braby, M. F. , Heath, A. , Lohman, D. J. , Mathew, J. , Rand, D. B. & Travassos, M. A. (2002). The ecology and evolution of ant association in the Lycaenidae (Lepidoptera). Annual Review of Entomology 47(1), 733–771. [DOI] [PubMed] [Google Scholar]
- Polgar, C. A. , Primack, R. B. , Williams, E. H. , Stichter, S. & Hitchcock, C. (2013). Climate effects on the flight period of Lycaenid butterflies in Massachusetts. Biological Conservation 160, 25–31. [Google Scholar]
- Posledovich, D. , Toftegaard, T. , Wiklund, C. , Ehrlén, J. & Gotthard, K. (2018). Phenological synchrony between a butterfly and its host plants: experimental test of effects of spring temperature. The Journal of Animal Ecology 87(1), 150–161. [DOI] [PubMed] [Google Scholar]
- Pounds, J. A. , Fogden, M. P. L. & Campbell, J. H. (1999). Biological response to climate change on a tropical mountain. Nature 398(6728), 611–615. [Google Scholar]
- Pöyry, J. , Leinonen, R. , Söderman, G. , Nieminen, M. , Heikkinen, R. K. & Carter, T. R. (2011). Climate‐induced increase of moth multivoltinism in boreal regions. Global Ecology and Biogeography 20(2), 289–298. [Google Scholar]
- Pöyry, J. , Luoto, M. , Heikkinen, R. K. , Kuussaari, M. & Saarinen, K. (2009). Species traits explain recent range shifts of Finnish butterflies. Global Change Biology 15(3), 732–743. [Google Scholar]
- Rafferty, N. E. , CaraDonna, P. J. & Bronstein, J. L. (2015). Phenological shifts and the fate of mutualisms. Oikos 124(1), 14–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruf, C. & Fiedler, K. (2002). Tent‐based thermoregulation in social caterpillars of Eriogaster lanestris (Lepidoptera: Lasiocampidae): behavioral mechanisms and physical features of the tent. Journal of Thermal Biology 27(6), 493–501. [Google Scholar]
- Saastamoinen, M. & Hanski, I. (2008). Genotypic and environmental effects on flight activity and oviposition in the Glanville fritillary butterfly. The American Naturalist 171(6), 701–712. [DOI] [PubMed] [Google Scholar]
- Saastamoinen, M. , Ikonen, S. & Hanski, I. (2009). Significant effects of Pgi genotype and body reserves on lifespan in the Glanville fritillary butterfly. Proceedings of the Royal Society B: Biological Sciences 276(1660), 1313–1322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheffers, B. R. , De Meester, L. , Bridge, T. C. L. , Hoffmann, A. A. , Pandolfi, J. M. , Corlett, R. T. , Butchart, S. H. M. , Pearce‐Kelly, P. , Kovacs, K. M. , Dudgeon, D. , Pacifici, M. , Rondinini, C. , Foden, W. B. , Martin, T. G. , Mora, C. , et al. (2016). The broad footprint of climate change from genes to biomes to people. Science 354(6313), 719. [DOI] [PubMed] [Google Scholar]
- Schneider, S. H. & Root, T. L. (2002). Wildlife Responses to Climate Change: North American Case Studies. Island Press, Washington, DC. [Google Scholar]
- Schweiger, O. , Settele, J. , Kudrna, O. , Klotz, S. & Kühn, I. (2008). Climate change can cause spatial mismatch of trophically interacting species. Ecology 89(12), 3472–3479. [DOI] [PubMed] [Google Scholar]
- Scriber, J. M. (2020). Assessing ecological and physiological costs of melanism in North American Papilio glaucus females: two decades of dark morph frequency declines. Insect Science 27(3), 583–612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seebacher, F. , White, C. R. & Franklin, C. E. (2015). Physiological plasticity increases resilience of ectothermic animals to climate change. Nature Climate Change 5(1), 61–66. [Google Scholar]
- Selwood, K. E. , McGeoch, M. A. & MacNally, R. (2015). The effects of climate change and land‐use change on demographic rates and population viability. Biological Reviews 90(3), 837–853. [DOI] [PubMed] [Google Scholar]
- Sheridan, J. A. & Bickford, D. (2011). Shrinking body size as an ecological response to climate change. Nature Climate Change 1(8), 401–406. [Google Scholar]
- Singer, M. C. & Parmesan, C. (2010). Phenological asynchrony between herbivorous insects and their hosts: signal of climate change or pre‐existing adaptive strategy? Philosophical Transactions of the Royal Society B: Biological Sciences 365(1555), 3161–3176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soroye, P. , Newbold, T. & Kerr, J. (2020). Climate change contributes to widespread declines among bumble bees across continents. Science 367(6478), 685–688. [DOI] [PubMed] [Google Scholar]
- Sparks, T. H. , Roy, D. B. & Dennis, R. L. H. (2005). The influence of temperature on migration of Lepidoptera into Britain. Global Change Biology 11(3), 507–514. [Google Scholar]
- Stålhandske, S. , Olofsson, M. , Gotthard, K. , Ehrlén, J. , Wiklund, C. & Leimar, O. (2016). Phenological matching rather than genetic variation in host preference underlies geographical variation in host plants used by orange tip butterflies. Biological Journal of the Linnean Society 119, 1060–1067. [Google Scholar]
- Stireman, J. O. , Dyer, L. A. , Janzen, D. H. , Singer, M. S. , Lill, J. T. , Marquis, R. J. , Ricklefs, R. E. , Gentry, G. L. , Hallwachs, W. , Coley, P. D. , Barone, J. A. , Greeney, H. F. , Connahs, H. , Barbosa, P. , Morais, H. C. , et al. (2005). Climatic unpredictability and parasitism of caterpillars: implications of global warming. Proceedings of the National Academy of Sciences of the United States of America 102(48), 17384–17387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stork, N. E. (2018). How many species of insects and other terrestrial arthropods are there on Earth? Annual Review of Entomology 63, 31–45. [DOI] [PubMed] [Google Scholar]
- Thomas, C. D. , Franco, A. M. A. & Hill, J. K. (2006). Range retractions and extinction in the face of climate warming. Trends in Ecology & Evolution 21(8), 415–416. [DOI] [PubMed] [Google Scholar]
- Thomas, J. A. (2005). Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups. Philosophical Transactions of the Royal Society B: Biological Sciences 360(1454), 339–357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas, J. A. (2016). Butterfly communities under threat. Science 353(6296), 216–218. [DOI] [PubMed] [Google Scholar]
- Van Der Kolk, H. J. , Wallis DeVries, M. F. & Van Vliet, A. J. H. (2016). Using a phenological network to assess weather influences on first appearance of butterflies in the Netherlands. Ecological Indicators 69, 205–212. [Google Scholar]
- Van Dyck, H. , Bonte, D. , Puls, R. , Gotthard, K. & Maes, D. (2015). The lost generation hypothesis: could climate change drive ectotherms into a developmental trap? Oikos 124(1), 54–61. [Google Scholar]
- Van Nouhuys, S. & Lei, G. (2004). Parasitoid‐host metapopulation dynamics: the causes and consequences of phenological asynchrony. Journal of Animal Ecology 73(3), 526–535. [Google Scholar]
- Vandergast, A. G. , Bohonak, A. J. , Hathaway, S. A. , Boys, J. & Fisher, R. N. (2008). Are hotspots of evolutionary potential adequately protected in southern California? Biological Conservation 141(6), 1648–1664. [Google Scholar]
- Varshney, R. K. , Thudi, M. , Pandey, M. K. , Tardieu, F. , Ojiewo, C. , Vadez, V. , Whitbread, A. M. , Siddique, K. H. , Nguyen, H. T. , Carberry, P. S. & Bergvinson, D. (2018). Accelerating genetic gains in legumes for the development of prosperous smallholder agriculture: integrating genomics, phenotyping, systems modelling and agronomy. Journal of Experimental Botany 69(13), 3293–3312. [DOI] [PubMed] [Google Scholar]
- Visser, M. E. , Bath, C. & Lambrechts, M. M. (2004). Global climate change leads to mistimed avian reproduction. Advances in Ecological Research 2504(04), 89–110. [Google Scholar]
- Visser, M. E. , Holleman, L. J. M. & Gienapp, P. (2006). Shifts in caterpillar biomass phenology due to climate change and its impact on the breeding biology of an insectivorous bird. Oecologia 147(1), 164–172. [DOI] [PubMed] [Google Scholar]
- Wagner, D. L. (2020). Insect declines in the Anthropocene. Annual Review of Entomology 65, 457–480. [DOI] [PubMed] [Google Scholar]
- Wang, L. , Yang, S. , Zhao, K. & Han, L. (2015). Expression profiles of the heat shock protein 70 gene in response to heat stress in Agrotis c‐nigrum (Lepidoptera: Noctuidae). Journal of Insect Science 15(1), 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warren, M. S. , Hill, J. K. , Thomas, J. A. , Asher, J. , Fox, R. , Huntley, B. , Roy, D. B. , Willis, S. G. & Greatorex‐Davies, J. N. (2001). Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 414(6859), 65–69. [DOI] [PubMed] [Google Scholar]
- Warren, R. J. & Bradford, M. A. (2014). Mutualism fails when climate response differs between interacting species. Global Change Biology 20(2), 466–474. [DOI] [PubMed] [Google Scholar]
- Watt, W. B. (1983). Adaptation at specific loci. II. Demographic and biochemical elements in the maintenance of the Colias PGI polymorphism. Genetics 103(4), 691–724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watt, W. B. (1992). Eggs, enzymes, and evolution: natural genetic variants change insect fecundity (allozymes/fitness components/global warming/phosphoglucose isomerase/thermal ecology). Proceedings of the National Academy of Sciences of the United States of America 89(22), 10608–10612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss, S. B. , Murphy, D. D. & White, R. R. (1988). Sun, slope, and butterflies: topographic determinants of habitat quality for Euphydryas editha . Ecology 69(5), 1486–1496. [Google Scholar]
- Wepprich, T. , Adrion, J. R. , Ries, L. , Weidmann, J. & Haddad, N. (2019). Butterfly abundance declines over 20 years of systematic monitoring in Ohio, USA. PLoS One 14(7), e0216270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheat, C. W. (2010). Phosphoglucose isomerase (Pgi) performance and fitness effects among arthropods and its potential role as an adaptive marker in conservation genetics. Conservation Genetics 11(2), 387–397. [Google Scholar]
- Williams, D. W. & Liebhold, A. M. (1995). Forest defoliators and climate change: potential changes in spatial distribution of outbreaks of western spruce budworm (Lepidoptera: Tortricidae) and gypsy moth (Lepidoptera: Lymantriidae). Environmental Entomology 24(1), 1–9. [Google Scholar]
- Willis, C. G. , Ruhfel, B. R. , Primack, R. B. , Miller‐Rushing, A. J. , Losos, J. B. & Davis, C. C. (2010). Favorable climate change response explains non‐native species' success in Thoreau's woods. PLoS One 5(1), e8878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson, R. J. , Brooks, S. J. & Fenberg, P. B. (2019). The influence of ecological and life history factors on ectothermic temperature–size responses: analysis of three Lycaenidae butterflies (Lepidoptera). Ecology and Evolution 9(18), 10305–10316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson, R. J. , Gutiérrez, D. , Gutiérrez, J. , Martínez, D. , Agudo, R. & Monserrat, V. J. (2005). Changes to the elevational limits and extent of species ranges associated with climate change. Ecology Letters 8(11), 1138–1146. [DOI] [PubMed] [Google Scholar]
- Wilson, R. J. & Maclean, I. M. D. (2011). Recent evidence for the climate change threat to Lepidoptera and other insects. Journal of Insect Conservation 15(1), 259–268. [Google Scholar]
- Winder, M. & Schindler, D. E. (2004). Climate change uncouples trophic interactions in an aquatic ecosystem. Ecology 85(8), 2100–2106. [Google Scholar]
- Wu, C. H. , Holloway, J. D. , Hill, J. K. , Thomas, C. D. , Chen, I. C. & Ho, C. K. (2019). Reduced body sizes in climate‐impacted Borneo moth assemblages are primarily explained by range shifts. Nature Communications 10, 4612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wymore, A. S. , Keeley, A. T. H. , Yturralde, K. M. , Schroer, M. L. , Propper, C. R. & Whitham, T. G. (2011). Genes to ecosystems: exploring the frontiers of ecology with one of the smallest biological units. New Phytologist 191(1), 19–36. [DOI] [PubMed] [Google Scholar]
- Xing, S. , Bonebrake, T. C. , Ashton, L. A. , Kitching, R. L. , Cao, M. , Sun, Z. , Ho, J. C. & Nakamura, A. (2018). Colors of night: climate–morphology relationships of geometrid moths along spatial gradients in southwestern China. Oecologia 188(2), 537–546. [DOI] [PubMed] [Google Scholar]
- Xing, S. , Bonebrake, T. C. , Tang, C. C. , Pickett, E. J. , Cheng, W. , Greenspan, S. E. , Williams, S. E. & Scheffers, B. R. (2016). Cool habitats support darker and bigger butterflies in Australian tropical forests. Ecology and Evolution 6(22), 8062–8074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, S. , Han, L. , Zhao, K. & Ye, L. (2015). Expression profile of two HSP70 chaperone proteins in response to extreme thermal acclimation in Xestia c‐nigrum (Lepidoptera: Noctuidae). Florida Entomologist 98(2), 506–515. [Google Scholar]
- Zera, A. J. & Denno, R. F. (1997). Physiology and ecology of dispersal polymorphism in insects. Annual Review on Entomology 42, 207–230. [DOI] [PubMed] [Google Scholar]
- Zeuss, D. , Brandl, R. , Brändle, M. , Rahbek, C. & Brunzel, S. (2014). Global warming favours light‐coloured insects in Europe. Nature Communications 5, 3874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu, K. , Woodall, C. W. & Clark, J. S. (2012). Failure to migrate: lack of tree range expansion in response to climate change. Global Change Biology 18(3), 1042–1052. [Google Scholar]