
Figure 3. Mitochondria control alternative splicing in roots through TOR kinase activity.
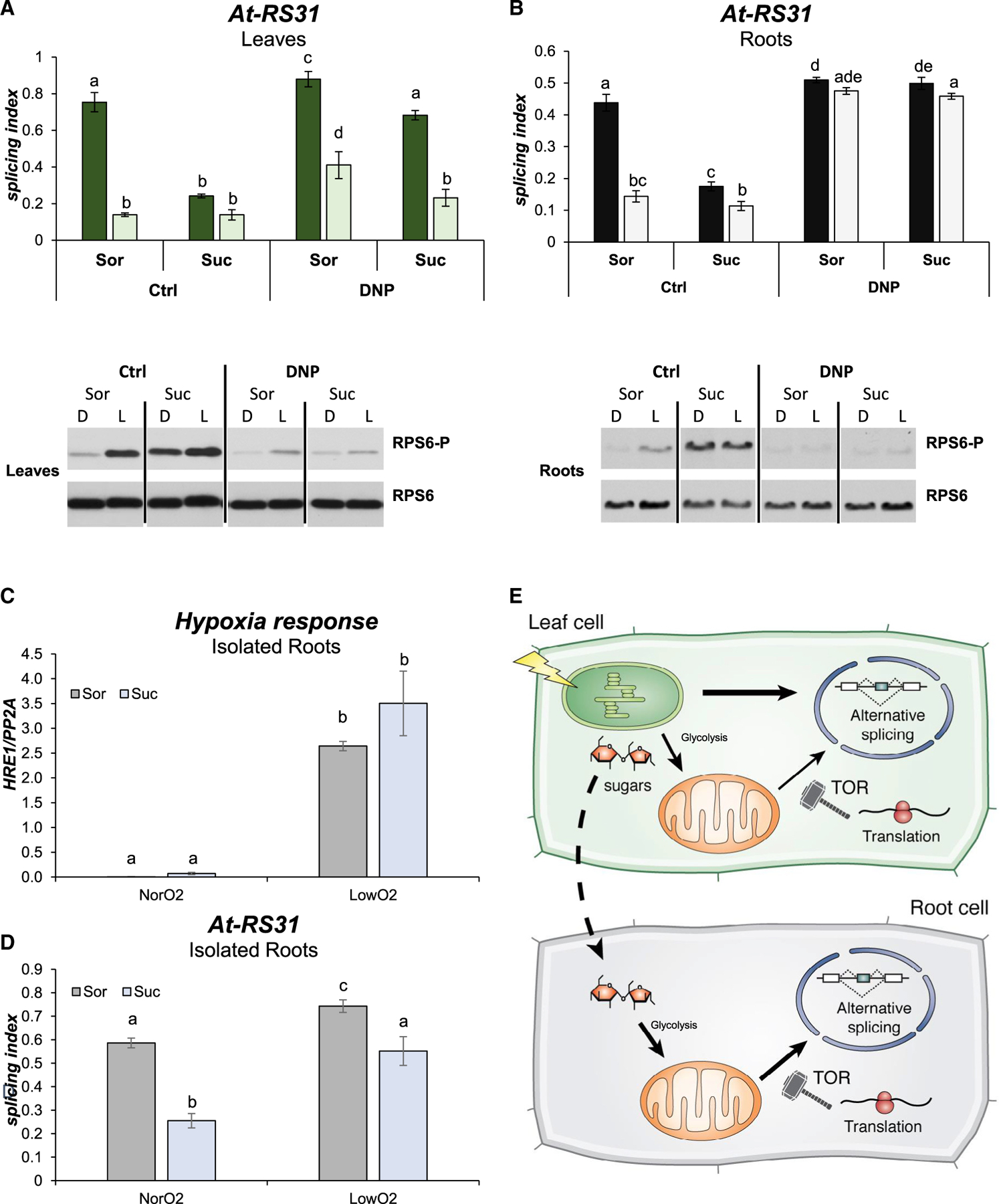
(A and B) Disrupting proton gradients abolishes light/dark splicing changes and TOR kinase activity modulation in roots. The ionophore 2,4-dinitrophenol (DNP) was used in a 20-μM concentration to incubate plants under the light/dark protocol. Suc, sucrose 100 mM. Sor, sorbitol 100 mM as osmotic control. Splicing indexes of At-RS31 are shown for leaves (A) and roots (B) in the top panels. Lighter bars, light; darker bars, darkness. Data represent splicing index means ± standard error (n = 4). The same letters indicate means that are not statistically different (p > 0.05). Bottom panels are western blot images of RPS6 phosphorylation (RPS6-P) and total RPS6 levels in leaves (A) and in roots (B).
(C and D) Low oxygen levels cause reduced alternative splicing responses. Plants were grown in constant light for 2 weeks, transferred to darkness for 48 h. Seedlings were dissected and isolated roots were treated with 50 mM sucrose or sorbitol in the presence of low oxygen (LowO2) or air (NorO2). Data represent splicing index means ± standard errors (n = 4). The same letters indicate means that are not statistically different (p > 0.05).
(C) HRE1 (hypoxia-inducible ethylene response factor 1) expression.
(D) At-RS31 alternative splicing index.
(E) Working model: different retrograde signals regulate alternative splicing through inter-organellar communication. Photosynthesis in the chloroplasts generates different signals that modulate alternative splicing in the nucleus of photosynthetic leaf cells. Among these signals, sucrose is also transported to the roots and feeds the mitochondria after glycolysis and activates TOR, thus changing alternative splicing in this heterotrophic tissue. Art by Dr. Luciana Giono.