

Summary
Fibroblasts are diverse mesenchymal cells that participate in tissue homeostasis and disease by producing complex extracellular matrix and creating signaling niches through biophysical and biochemical cues. Transcriptionally and functionally heterogeneous across and within organs, fibroblasts encode regional positional information and maintain distinct cellular progeny. We summarize their development, lineages, functions, and contributions to fibrosis in four fibroblast-rich organs: skin, lung, skeletal muscle, and heart. We propose that fibroblasts are uniquely poised for tissue repair by easily reentering the cell cycle and exhibiting a reversible plasticity in phenotype and cell fate. These properties, when aberrantly activated, drive fibrotic disorders in humans.
Horsley eTOC in brief
A detailed mechanistic, molecular and functional view of the commonalities and organ-specific features of fibroblasts in both health and disease is just beginning to emerge.
Introduction
Fibroblasts are canonically referred to as cells that create and maintain anatomically diverse array of extracellular matrix (ECM)-rich connective tissues to support a broad range of essential organ functions, like resistance to blunt and sharp injuries in the skin or organ-wide stretching and elastic recoiling in the intact breathing lung. In doing so, fibroblasts provide essential niches and positional information for neighboring cells via microarchitectural, biomechanical, and biochemical cues in the ECM and the regulated secretion of soluble mediators such as cytokines, growth factors and metabolites (Figure 1). Beyond producing connective tissues, fibroblasts serve as the progenitors for specialized mesenchymal cell types, such as for bone-forming osteoblasts or lipid-filled adipocytes during embryonic development, adult homeostasis and during injury, repair, and remodeling. In this review, we use the term “fibroblasts” to refer to cells that: 1) secrete many of the same structural and signaling macromolecules that contribute to tissue’s extracellular space, 2) adopt a transient and contractile myofibroblast phenotype in response to tissue damage, 3) act as signaling niche cells for tissue-resident stem cells, and/or 4) serve as progenitors, sometimes called mesenchymal stem cells, for specialized differentiated mesenchymal cells (Lemos and Duffield, 2018; Pittenger et al., 2019; Pittenger et al., 1999).
Figure 1. Summary of fibroblast outputs and functions.
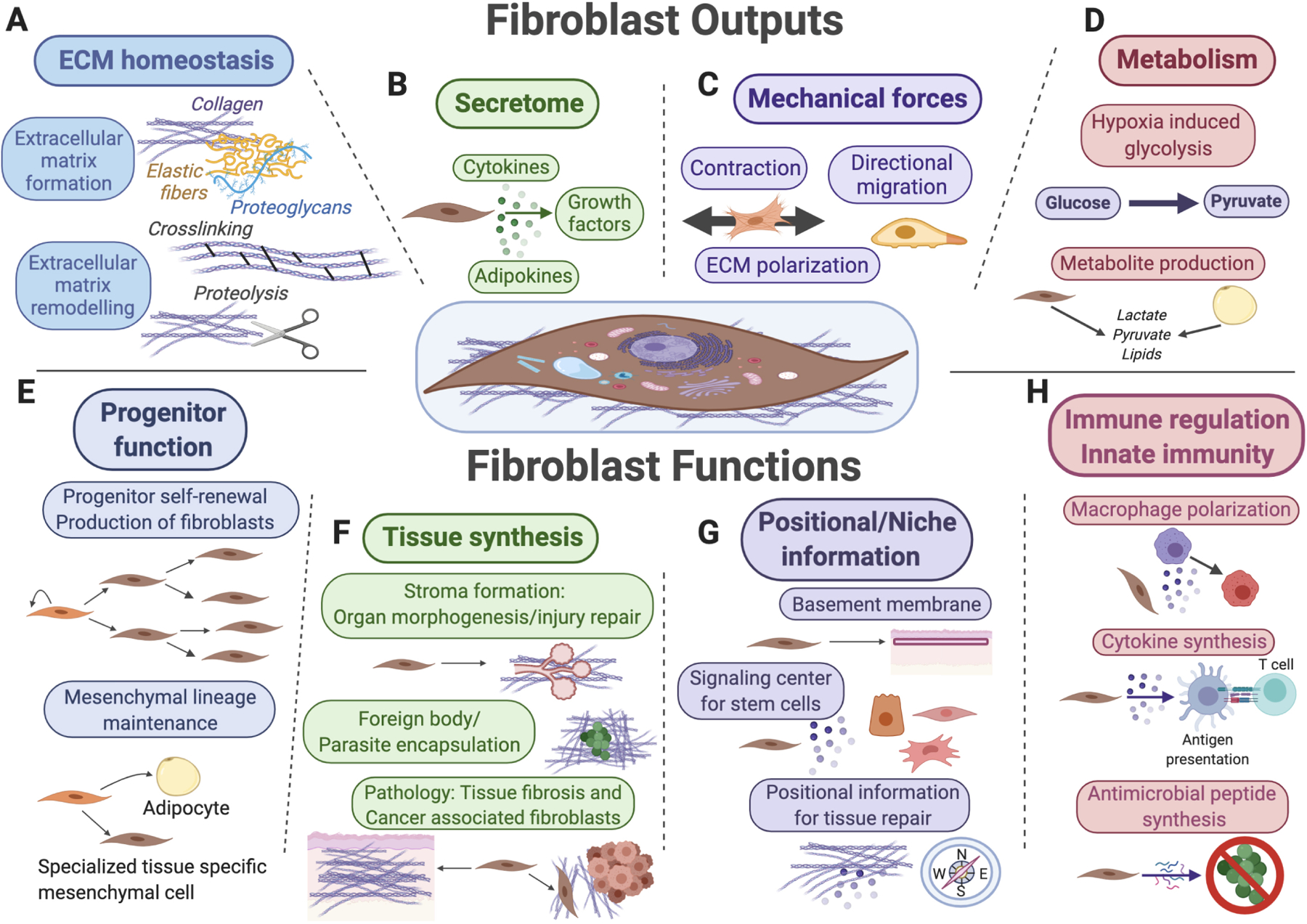
Key functions for fibroblasts (shown in the center) and their mesenchymal lineages include extracellular matrix (ECM) secretion and remodeling (A), secretion of signaling factors for surrounding cells (B), mechanical force generation (C), and regulation of tissue metabolism and metabolite secretion (D). Fibroblasts also function as progenitor cells for mesenchymal lineages (E), as “makers” of new tissue during organ morphogenesis, tissue repair and upon various pathological conditions (F), as sources of positional information across distinct anatomical regions of the same organ and as key signal contributors toward stem cell niches (G), as well as target cells and reciprocal modulators of diverse innate and adaptive immune functions (H).
Fibroblasts were first described as a distinct cell type in 1858 by German pathologist Rudolf Virchow, who called them “Spindelzelle des Bindegewebes” – “spindle-shaped cells of the connective tissue” (Virchow, 1858) (Figure 2A). The term “fibroblast” was first proposed by Ernst Ziegler to describe cells that produce new connective tissue upon healing (Ziegler, 1895) (Figure 2B) and this observation was replicated by Santiago Ramón y Cajal who observed “célula fusiforme” or “fibro-células” as essential producers of granulation tissue in both healing skin wounds and scars (Cajal, 1896) (Figure 2C).
Figure 2. History of fibroblast discovery.
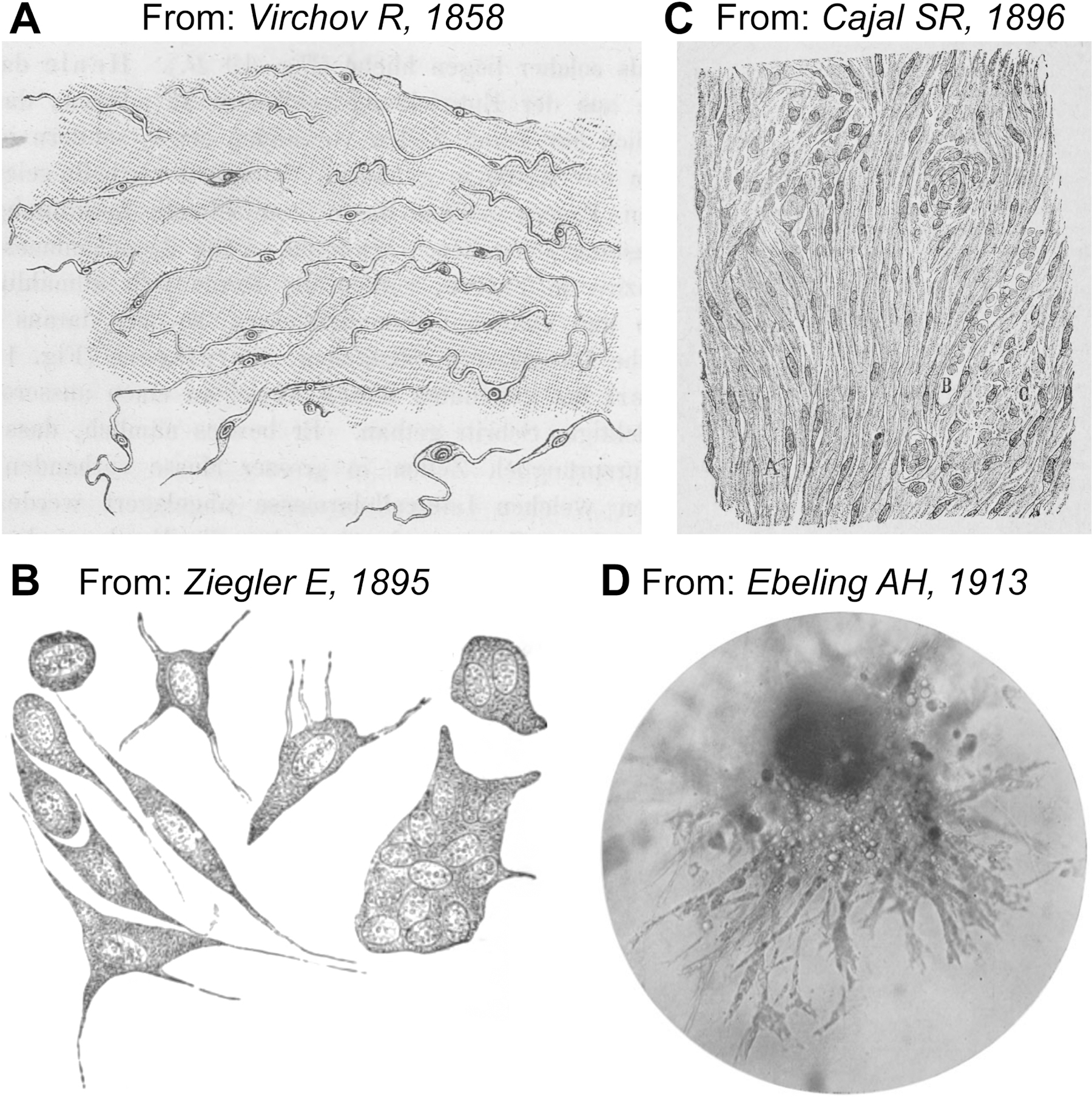
A. First drawing of fibroblasts by Rudolf Virchow as “spindle-shaped” cells embedded within the connective tissue of pig embryo. Modified from Virchow (1858). B. Drawing by Ernst Ziegler, who first proposed the term “fibroblast” to describe cells that produce new connective tissue upon healing. Various forms of cells in the new granulation tissue are shown. Mononuclear fibroblast-shaped cells are in the bottom left corner. Modified from Ziegler (1895). C. Drawing of fibroblasts as fusiform cells within newly formed connective tissue of a “painful” keloid. Modified from Cajal (1896). D. Microphotograph of fibroblasts established from embryonic chick heart explant. Cells after 75 passages are shown. Modified from Ebeling (1913).
Fibroblast research was facilitated by the advent of in vitro techniques developed in the 1900s. These methods allowed culture of primary fibroblasts derived from embryonic chick heart explants, which could be easily propagated upon passaging before other cell types (Burrows and Neymann, 1917; Carrel, 1912; Ebeling, 1913; Hogue, 1919) (Figure 2D). The establishment of the 3T3 fibroblast cell line derived from mouse embryos more than fifty years later (Todaro and Green, 1963; Todaro et al., 1964) further advanced our understanding of the biology and lineage potential of fibroblasts. These discoveries include, among others, the identification of fibroblast growth factors (FGFs) (Gospodarowicz, 1974), the phenomenon of multi-lineage differentiation of cultured fibroblasts into bone, cartilage, and adipose cells (Junker et al., 2010), and the key role of fibroblasts in the production, remodeling, and contraction of ECM.
Despite the large number of in vitro fibroblast studies, the in vivo relevance of these cell culture observations remains unclear, and the true spectrum of fibroblasts’ properties and their lineage potential in their native in vivo environment is only recently being explored in depth. This is largely enabled by the advent of new genetic model organisms that permit specific labeling, tracing and mutating of fibroblasts in tissues and by the application of single-cell genomic technologies. Emerging evidence implicates the role of fibroblasts in driving significant changes in the tumor-associated macroenvironment as have been extensively reviewed elsewhere (Sahai et al., 2020). Here, we focus on fibroblast development and their role in adult tissue homeostasis and repair including recent evidence that highlights the function and heterogeneity of fibroblasts across and within organs and how they are regulated at the molecular level to contribute to tissue development, homeostasis, repair and fibrotic diseases.
Cross organ commonalities in fibroblast biology
Single-cell RNA-sequencing (scRNA-seq) data from several organs has revealed an unappreciated degree of heterogeneity in fibroblasts within and across tissues. Comparing fibroblasts from mouse heart, skeletal muscle, intestine and bladder revealed that less than 20% of fibroblast-enriched genes overlapped between these four organs (Muhl et al., 2020). Most recently, an even broader cross-tissue comparison of mouse scRNA-seq data identified two universal fibroblast populations, expressing peptidase inhibitor Pi16 or collagen Col15a1 each with shared enrichment for the ECM factor dermatopontin (Dpt). The latter gene marks the majority of PDGFRα+ cells in the surveyed tissues by genetic lineage tracing in mice (Buechler et al., 2021). Murine fibroblast populations expressing these universal markers persisted throughout injury, tumorigenesis, or inflammation with additional specialized subpopulations emerging in these perturbed states. Notably, a population similar to Pi16+ mouse fibroblasts was also found in unperturbed human tissues (Buechler et al., 2021), suggesting a cross species contribution to homeostasis. Further analysis will be required to determine the convergence and divergence of fibroblast populations across murine organs, especially since not all fibroblasts express Pdgfra, such as lipofibroblasts in the lung (Xie et al., 2018) or skin fibroblasts after wounding (Guerrero-Juarez et al., 2019), which also show largely nonoverlapping patterns of Dpt and Col15a1 and nearly absent Pi16 expression. Beyond gene markers, functional properties are the ultimate discriminator of shared vs. unique themes in fibroblast biology. In this section, we outline the shared functions of fibroblasts across multiple tissues and, in a separate section, highlight numerous tissue-specific differences.
Scaffold and signaling: Common fibroblast functions
A major shared function of fibroblasts is ECM synthesis to create connective tissue by depositing fiber- and sheet-forming collagens, proteoglycans, elastin, fibronectin, microfibrillar proteins and laminins, that collectively comprise the “matrisome”. Fibroblasts also actively remodel ECM microstructure through covalent crosslinking, protein glycosylation and controlled proteolysis via balanced secretion of modifying enzymes such as lysyl oxidase, matrix metalloproteinases (MMPs), and MMP inhibitors (Lu et al., 2011). The ratio of specific ECM molecules and nuanced remodeling activity by fibroblasts can produce an array of compositionally, micro-anatomically, biomechanically, and functionally distinct ECM across organs that can support a range of specialized cells, such as keratinocytes in the resilient and soft skin, epithelial cells in the malleable and elastic lung, striated muscle fibers in the contractile skeletal muscle, as well as endothelial cells of blood vessels (Figure 1A) (Hynes and Naba, 2012). Fibroblasts also “tug and pull” on their ECM, resulting in tissue-level mechanical forces and matrix polarization (Huang et al., 2012) (Figure 1C) and contribute to tissue specific matrisomes and tissue mechanics.
Beyond the matrisome, fibroblasts secrete numerous cytokines, adipokines and growth factors (Figure 1B, 1H) whose properties, including diffusion dynamics, are modulated by the ECM, and converge to create signaling niches and positional cues for diverse other cells, including but not limited to tissue-resident stem cells and immune cells (McGee et al., 2013) (Figure 1G). In this light, fibroblasts’ role in encoding positional information for other cells is particularly important for proper embryonic development, as revealed in classic tissue recombination studies. For example, when embryonic chick skin epithelium and mesenchyme from scale or feather producing body regions were exchanged, the type of skin appendage that formed was instructed by fibroblast-containing dermis (Dhouailly and Sengel, 1975).
Signaling between tissue resident cells and fibroblasts can establish regional differences and lineage trajectories of fibroblasts in tissues. For instance, synovial fibroblasts exhibit positional identity that is induced by endothelium-derived Notch signaling (Wei et al., 2020). Another mechanism by which instructive differences in fibroblast biology are established is through expression of specific Homeobox (HOX) transcription factors, which specify body plan along major axes – cranio-caudal, dorsal-ventral and proximal-distal directions, including in humans (Chang et al., 2002). For example, HOXA13 regulates distal identity and is expressed specifically by finger and foreskin human fibroblasts (Rinn et al., 2006). Regional expression differences exist in other genes, that are likely downstream of HOX factors, such as Agouti in mice (Candille et al., 2004), Wingless-related integration site (WNT) pathway antagonist DKK1 in humans (Yamaguchi et al., 2004), and fibronectin in both rodents and humans (Yasuda et al., 2006). Therefore, distinct “HOX codes” of skin fibroblasts drive regional differences in matrisome and signaling factors that, in turn, serve as “information” for adjacent cell types. Reestablishing this positional information may also be an important step during tissue regeneration after injury. For example, restoration of fibroblast HOX codes may function as a pioneering event to drive skin regeneration in adult mice (Abbasi et al., 2020). Beyond skin, positionally distinct fibroblasts with unique HOX codes have been recently identified in synovial joints in mice and humans (Frank-Bertoncelj et al., 2017).
Myofibroblasts: Contraction and coordination of tissue repair
In adult organs, fibroblasts are relatively quiescent unless tissue repair mechanisms or dynamic structural changes are initiated. Recent single-cell analyses indicate that during tissue development and repair, fibroblasts display transcriptional changes similar to cellular differentiation trajectories, suggesting an adherence to lineage hierarchy (Abbasi et al., 2020; Guerrero-Juarez et al., 2019; Phan et al., 2020; Phan et al., 2021). Quiescent fibroblasts have also been shown to function as progenitors that can be induced to rapidly divide to produce many more ECM-secreting fibroblasts and additional distinct mesenchymal lineages, such as adipocytes, in response to injury and hair cycling in the skin (Figures 3, 4) (Junker et al., 2010; Rivera-Gonzalez et al., 2016).
Figure 3. Key roles of PDGF, TGFβ and WNT signaling pathways in regulating fibroblast functions.
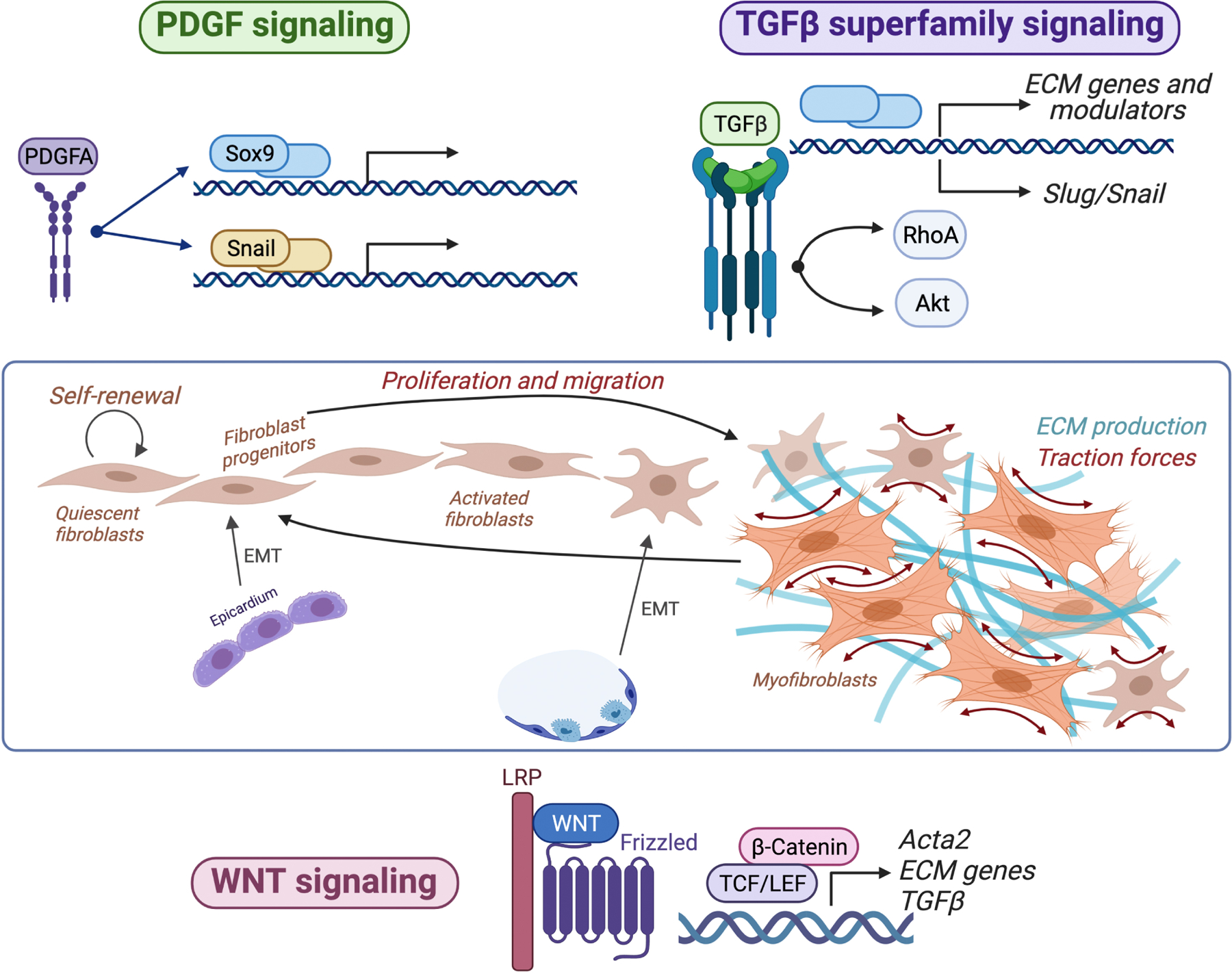
Platelet derived growth factor (PDGF) signaling (blue) regulates diverse aspects of fibroblast development and homeostasis, including epithelial-to-mesenchymal transition (EMT) in the embryonic heart, long-term self-renewal, and proliferation in adult tissues. Signaling via the transforming growth factor β (TGFβ) superfamily of ligands promotes myofibroblast state activation, including contractile protein and extracellular matrix (ECM) gene expression. Among other effects, TGFβ superfamily signaling can induce fibroblast proliferation and lineage transitions by other cells toward a fibroblast state, including via EMT in the lung. WNT signaling regulates fibroblast proliferation, migration, myofibroblast state activation, and ECM deposition. All three pathways can activate transcription of genes to control fibroblast biology and TGFβ activates Akt and RhoA to induce cellular contraction.
Figure 4. Diverse cellular sources of myofibroblasts.
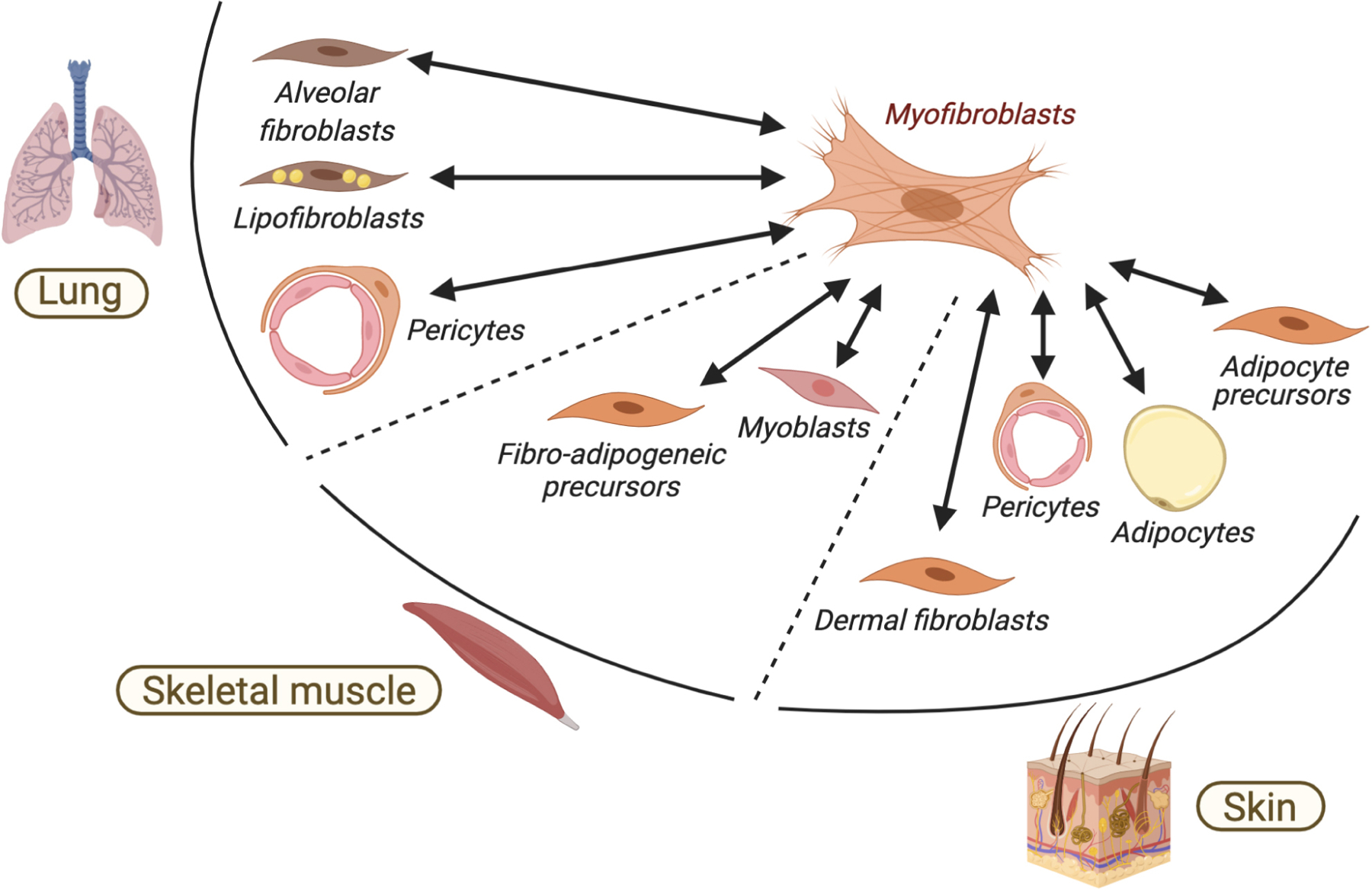
Diverse tissue resident mesenchymal cells, including specialized fibroblast progenitors, pericytes and adipocytes can become activated and undergo reversible conversion toward a myofibroblast state. Examples of lung, skeletal muscle and skin are provided.
To facilitate tissue repair, signaling and physical factors induce quiescent fibroblasts to form myofibroblasts, which through their expression of contractile proteins such as αSMA orchestrate biomechanical remodeling and contraction via traction force (Pakshir et al., 2019) onto the voluminous amounts of new ECM that they themselves secrete (Hinz, 2010; Tschumperlin, 2013) (Figure 3). Compared to quiescent fibroblasts, mature myofibroblasts adhere to ECM for longer (Hinz, 2007) even in the presence of the same signals, suggesting that disengagement of focal adhesions may be essential for myofibroblast state termination (Thannickal, 2013). External adhesion “information” is transduced in myofibroblasts via the cytoskeleton and involves activation of tension sensitive myocardin-related transcription factor A (MRTF-A) and serum response factor (SRF) (Crider et al., 2011), among other mechanisms.
Beyond contraction, myofibroblasts also function to transform the surrounding tissue environment by modulating resident immune cell functions (Ferrer et al., 2017; Van Linthout et al., 2014) and phagocytosing dead cells (Nakaya et al., 2017). While myofibroblast functions are vital during acute injury repair, their aberrant activation upon chronic injury or sustained inflammation can lead to disorganized and excessive ECM production, promoting localized scarring, diffuse fibrosis and, in some instances, aiding in tumorigenesis (Desmouliere et al., 2005; Shi-wen et al., 2009). This excessively stiff and compositionally abnormal ECM disrupts microarchitecture, results in loss of other tissue-resident cells, causing organ dysfunction that can range from mild derangements to catastrophic failure. Fibrosis is estimated to contribute to almost 50% of all deaths in the developed world (Friedman et al., 2013) and despite the existence of drugs that can delay its progression, to date there exists no truly curative treatment (Dempsey et al., 2019). Thus, illuminating the fibrogenic functions of tissue fibroblasts holds remarkable therapeutic potential, which we discuss below.
Myofibroblasts arise in response to several signaling pathways including transforming growth factor β (TGFβ), WNT, and PDGF signaling and to some extent by inflammatory cytokines, such as tumor necrosis factor α (TNFα), interleukin (IL)-1 or IL-6 (Figure 3). These factors act on the quiescent, tissue-resident fibroblasts, which are viewed as the dominant source of myofibroblasts after injury. Of note, other specialized mesenchymal cells can also alter their lineage specificity and generate myofibroblasts (Figure 4). For example, pericytes, mesenchymal cells that surround blood vessels and that are transcriptionally distinct from fibroblast populations (Armulik et al., 2011), can migrate out of their perivascular location, develop myofibroblast properties and contribute to excessive ECM deposition in mouse models (Hung et al., 2013; Kuppe et al., 2020; Sava et al., 2017). A relatively small portion of myofibroblasts under certain wounding conditions in the skin and other organs has also been shown to derive from circulating hematopoietic progenitors, commonly myeloid cells (Guerrero-Juarez et al., 2019; Opalenik and Davidson, 2005; Sinha et al., 2018). Studies in mice have also revealed that mature adipocytes in the skin can deplete their lipid stores and convert into contractile myofibroblasts (Kruglikov and Scherer, 2017; Marangoni et al., 2015; Shook et al., 2020). Myoblasts in murine skeletal muscles can also form myofibroblasts as part of the persistent injury-repair cycle upon muscle dystrophy (Biressi et al., 2014). Finally, lipofibroblasts in the mouse lung can adopt functions of myofibroblasts including αSMA expression and ECM overproduction (El Agha et al., 2017; Rock et al., 2011).
While traditionally myofibroblasts were thought to be terminally differentiated, accumulating evidence indicates that they are, in fact, temporary and reversible (Hinz et al., 2012). Despite the prominent transcriptional and epigenetic changes that accompany myofibroblast formation, contractile gene expression and function by myofibroblasts decrease and eventually cease when tension reduces in late-stage skin wounds (Hinz et al., 2001). Moreover, myofibroblasts can display broader lineage plasticity and convert into other specialized mesenchymal lineages upon injury resolution. For example, myofibroblasts in large murine skin wounds terminate their contractile behavior and reprogram into new lipid-filled adipocytes in response to bone morphogenetic protein (BMP) ligands secreted by hair follicles (Plikus et al., 2017). During trauma-induced heterotopic ossification, fibroblasts can assume chondrogenic and osteogenic mesenchymal fates in extra-skeletal locations, such as in the skin, both in mice and humans (Cappato et al., 2020). Whether such heterogeneity in origin and function occurs in humans remains an active area of investigation.
Fibroblast plasticity
It is, thus, emerging that fibroblast lineages are plastic and do not follow the canonical stem cell lineage model, in which undifferentiated and long-term self-renewing progenitors unidirectionally produce differentiated and, typically, postmitotic progeny (Morrison and Spradling, 2008). Lineage plasticity displayed by fibroblasts during tissue repair resonates with that by other cell types, such as epithelial hair follicle stem cells. The latter normally produce hair-fated progeny but respond to wounding by temporarily switching to making short-lived epidermal keratinocytes to close the breached skin barrier (Ge et al., 2017; Ito et al., 2005). The crucial need to repair damaged tissue induces normally quiescent and, in some instances, postmitotic mesenchymal cells, such as adipocytes, to reenter cell cycle and produce new ECM-making and contracting myofibroblasts. Once the damage is healed, myofibroblasts revert to quiescence and in some instances prune via apoptosis or senescence (Demaria et al., 2014; Wilkinson et al., 2019). Fibroblast plasticity may be possible because they deposit most of their differentiated product (i.e. ECM proteins) outside of the cell (Hynes and Naba, 2012) as opposed to, for example, keratin filament-laden suprabasal epidermal cells or sarcomere protein-filled myofibers, which may permit fibroblasts to remain relatively unspecialized and exit cellular quiescence upon stimulation.
Epigenetic regulation of fibroblast identity and dynamic state transitions, including the myofibroblast state, is a subject of intense research. For example, several microRNAs can activate, sustain, and terminate the myofibroblastic state across different reparative contexts (Wei et al., 2019; Yang et al., 2013) and more durable epigenetic modifications have also been implicated (Duong and Hagood, 2018). For example, during non-scarring kidney repair, wound-responsive fibroblasts suppress RASAL1, a promoter that encodes an inhibitor of Ras oncoproteins, and this enables transient activation. RASAL1 suppression is reversible, and its eventual restoration is necessary for proper myofibroblast state resolution. Extended exposure to profibrotic signals, such as TGFβ, results in initial suppression, and subsequent hypermethylation of RASAL1 promoter, which causes irreversible transcriptional suppression, lasting myofibroblast activation and chronic fibrogenesis (Bechtel et al., 2010). It remains less clear which epigenome modifications in fibroblasts enable recapitulation of developmental programs during regenerative healing. Future work is needed to determine whether different fibroblast pools harbor distinct levels of latent plasticity, and whether fibroblasts retain an epigenetic memory of earlier reparative events that heightens their responsiveness to subsequent insults, as seen in epithelial cells (Naik et al., 2017). Answers to these questions will inform future therapeutic efforts at suppressing the dual nature of fibroblast plasticity in tissue repair, by restricting fibrogenic capacity in favor of readoption of native tissue functions.
Signaling regulation of fibroblast lineages and function
Key aspects of fibroblast biology, from proliferation and self-renewal to myofibroblast formation and differentiation to other mesenchymal lineages, are regulated by a diverse array of signaling pathways that act in autocrine, paracrine and endocrine manner. While many signaling factors can impact fibroblasts and contribute to fibrosis, such as FGFs (Xie et al., 2020), in this section, we review major roles played by platelet-derived growth factor (PDGF) pathway in fibroblast proliferation and self-renewal, TGFβ super family pathways in fibroblast function, canonical WNT pathway in fibroblast lineage specification and fibrosis, as well as mechanotransduction and danger-associated molecular pattern (DAMP) stimulation (Figure 3).
Role of PDGF signaling
Developmental expansion and long-term maintenance of fibroblast lineages requires their self-renewal via proliferation. This key property of fibroblasts critically relies on PDGF signaling. PDGF ligands act locally within tissues and can be produced by fibroblasts themselves or by other cell types to function as homo- and heterodimers of four different polypeptide chains, from A through D. While PDGF-AA, PDGF-AB and PDGF-BB are processed and secreted as dimers, PDGF-CC and PDGF-DD are secreted as inactive forms (Kazlauskas, 2017). PDGF ligands engage with PDGFRα and -β tyrosine kinase receptors, which activate multiple downstream signaling cascades, including the RAS/MAP kinase and AKT/PI3 kinase cascades, that in turn propagate signaling to their downstream effectors.
PDGFRα is commonly expressed by the progenitor cells of multiple mesenchymal lineages, including fibroblasts and adipocytes (Uezumi et al., 2014) and Pdgfra null mice die prior to birth, showing prominent deficiencies in cardiac mesoderm, somitic mesoderm, and neural crest derived mesenchyme (Soriano, 1997). In the developing mouse heart, PDGF signaling induces the expression of transcriptional regulators Sox9, Snail and Slug that induce epicardial cells to undergo EMT to form the majority of cardiac fibroblasts (Figure 3). Conditional gain- and loss-of-function studies in mice have illustrated the importance of PDGF signaling in maturation of postnatal fibroblast lineages, long-term maintenance of their progenitor function, and ECM homeostasis. For example, mice with constitutively active receptor Pdgfra display connective tissue hyperplasia and develop systemic fibrosis in multiple organs, including skin and heart (Gallini et al., 2016; Olson and Soriano, 2009). On the other hand, mice with Pdgfra deficiency display tissue hypoplasia. Likewise, tissue- and cell type-specific losses of Pdgfa ligand in mice lead to prominently reduced proliferation and self-renewing potential of dermal adipocyte precursors in the skin (Rivera-Gonzalez et al., 2016), αSMA+ dermal progenitors in the hair follicle (Gonzalez et al., 2017) and fibroblasts in the lung (Gouveia et al., 2018). Temporal regulation of PDGF activation during myocardial infarction induces cardiac fibroblast self-renewal and proliferation without triggering fibrosis (Asli et al., 2018) and pharmacological inhibition of PDGF signaling reduces ECM production during skeletal muscle repair (Smith et al., 2011). Thus, the PDGF signaling pathway serves as the major signaling regulator of fibroblast development and long-term homeostasis by supporting fibroblast stem cell self-renewal, proliferation, and migration.
Role of TGFβ superfamily signaling
During tissue repair, fibroblasts secrete and mechanically rearrange large quantities of new ECM. To perform these functions, fibroblasts must transition from a resting state, when ECM production is minimal, to a contractile myofibroblast state, when they upregulate matrisome synthesis and activate contractile apparatus. While the transition toward myofibroblast state is essential for tissue repair after injury, its aberrant and sustained switch critically drives fibrosis and contributes to cancer progression. Activation of myofibroblast state is dominantly controlled by TGFβ pathway (Massague, 2012) and human fibrotic tissues display elevated expression of TGFβ ligands in the lung, skin, and skeletal muscle (reviewed in Lodyga and Hinz, 2020).
Upon homeostasis, TGFβ ligands, together with latency-associated peptides (LAPs) and latent TGFβ-binding proteins (LTBPs) form large latent complexes (LLCs) (Shi et al., 2011), that are tethered in the ECM. Upon injury or fibrotic stimuli, TGFβs are rapidly released from the LLCs upon proteolytic cleavage of LAPs or via mechanical forces generated onto ECM. TGFβ ligands bind hetero-tetrameric receptor complexes and activate canonical SMAD pathway, including receptor-phosphorylated SMAD2/3, co-activator SMAD4, and inhibitory SMAD7 (Hata and Chen, 2016). Phosphorylated SMAD2/3 complexes translocate to the nucleus where they interact with other context-dependent transcription factors to regulate downstream genes (Verrecchia et al., 2001). TGFβ can also activate non-canonical signaling pathways implicated in fibrosis such as MAP kinases, Rho-like GTPase signaling, and phosphatidylinositol-3-kinase/AKT (Zhang, 2017). Because SMAD-driven mechanisms are the most well studied, they will be discussed in detail below.
Multiple studies reinforce the essential role of TGFβ signaling in ECM homeostasis, myofibroblast formation, and fibrosis development (Lodyga and Hinz, 2020). For instance, deletion of TGFβ receptor II in mouse fibroblasts abrogates contraction and ECM production in skin wounds (Denton et al., 2009). Postnatal activation of TGFβ receptor I in mouse fibroblasts recapitulates fibrotic phenotypes in the skin (Sonnylal et al., 2007) and heart even in the absence of injury (Nakajima et al., 2000). TGFβ-regulated SMAD2/3 complexes activate transcription of matrisome genes, contractile factors, such as αSMA, and connective tissue growth factor (CTCF), which also cooperates with TGFβ in to sustain a myofibroblast state (Tsai et al., 2018). Smad3 deletion in mice protects against bleomycin-induced lung fibrosis (Zhao et al., 2002), skin fibrosis (Lakos et al., 2004), and cardiac fibrosis (Dobaczewski et al., 2010). TGFβ signaling also impacts myofibroblast formation via metabolic reprogramming of fibroblasts through mitochondrial biogenesis or glycolysis (Bernard et al., 2015).
Other members of the TGFβ superfamily of ligands also impact fibroblast biology. BMP signaling, which activates SMAD1/5/8 proteins to alter gene expression with co-activator SMAD4 (von Bubnoff and Cho, 2001), impacts differentiation of several fibroblast-derived lineages (Wang et al., 2014). During regeneration of the skeletal muscle BMP signaling convert fibroblasts to myoblasts (de Lima et al., 2020) and during wound healing in the skin it reprograms myofibroblasts to adipocytes (Plikus et al., 2017). BMP is also required to maintain the specialized identity of the so-called dermal papilla (DP) fibroblasts that constitute essential signaling niche for epithelial stem cells in the hair follicle (Rendl et al., 2008).
Another TGFβ superfamily member, Activin A, which like TGFβ ligands activates SMAD2/3 (Pangas and Woodruff, 2000), is upregulated in human scars and other fibrotic diseases. When overexpressed from keratinocytes in mouse skin, Activin A upregulates gene expression within matrisome, secretome and modulating enzyme categories by skin fibroblasts, and reduces ECM deformability after injury (Wietecha et al., 2020 and references therein). Furthermore, Activin A inhibition can attenuate liver and lung fibrosis in mouse and rat models, respectively and may impact fibroblast biology in fibrosis more broadly (reviewed in Werner and Alzheimer, 2006). Thus, the TGFβ superfamily can activate distinct fibroblast mechanisms to drive tissue repair and contribute to fibrosis.
Role of WNT signaling
Canonical WNT pathway regulates fate specification of fibroblasts in development and can modulate the continuum of fibrosis and regeneration in adult tissues. Canonical WNT ligands bind to complexes of low-density lipoprotein receptors (LRPs) and Frizzled receptors on their target cells. In the absence of a WNT signal, cytoplasmic β-catenin is phosphorylated by a multiprotein destruction complex consisting of Axin, the adenomatous polyposis coli protein (APC), and several kinases, leading to its ubiquitin-mediated degradation. Upon ligand binding, the destruction complex is dissembled, leading to cytoplasmic β-catenin stabilization and translocation to the nucleus, where it forms complexes with transcription factors of the TCF/LEF family to regulate downstream gene expression (MacDonald et al., 2009).
During morphogenesis, canonical WNT pathway has been shown to regulate fate specification of fibroblast progenitors into various lineages, notably in the skin. WNT signaling is activated in the papillary fibroblast progenitors of the upper skin layer and then becomes progressively restricted to the so-called dermal condensate cells of developing hair follicles (DasGupta and Fuchs, 1999; Zhang et al., 2009). Dermal condensate cells eventually mature into adult DP fibroblasts, and this process fails in mice with the loss-of-function in WNT signaling (Millar, 2002). On the other hand, consistent with the inhibitory effect that WNT signaling exerts on adipocyte differentiation of mesenchymal cell in vitro, its activation in the so-called reticular fibroblasts of the lower skin layer inhibits formation of cutaneous adipocytes in development (Mastrogiannaki et al., 2016) and during regeneration in adult skin wounds (Plikus et al., 2017). Moreover, in skin wounds, transient WNT activity in myofibroblasts promotes regenerative healing with new hair follicles (Lim et al., 2018), while its chronic activity drives fibrotic response and failure to regenerate hairs (Gay et al., 2020). The impact of WNT signaling on fibroblast development and injury response in other tissues remains poorly understood.
Mechanotransduction signaling
Along with soluble growth factors, fibroblast behavior is prominently influenced by biochemical and biomechanical properties of the surrounding ECM, which vary between and even within the same organs. The biomechanical differences in ECM are sensed by fibroblasts through integrin-associated focal adhesions (Balaban et al., 2001) and are interpreted analogous to biochemical signals. Biomechanical cues from the ECM induce activation of myocardin-related transcription factors (Huang et al., 2012) and transcriptional cofactors YAP and TAZ (Jorgenson et al., 2017), as well as chromatin state changes (Le et al., 2016). Sensing of biomechanical inputs promotes fibroblast proliferation and can induce myofibroblast state (Huang et al., 2012) and acquisition of pro-fibrotic lineage identity (Mascharak et al., 2021), which in turn can contract ECM to release ECM-bound TGFβ (Wipff et al., 2007), further amplifying the response. Restoration of biomechanical tissue properties using biomaterials as well as direct pharmacological targeting of mechanosensitive pathways that can override this form of fibroblast activation are viewed as promising future antifibrotic strategies (Mascharak et al., 2021; Meli et al., 2020).
DAMP signaling
As key factors in tissue repair, fibroblasts are sensitive to an array of damage associated molecular pattern (DAMP) signals like intracellular macromolecules, including RNA, DNA, histones and heat shock proteins, released from damaged cells as well as to ECM molecule fragments (Turner, 2016). Sensing of DAMPs is mediated through activation of pattern recognition receptors (PRRs), such as transmembrane toll-like receptors (TLRs) and cytoplasmic nod-like receptors (NLRs) (Schaefer, 2014). While PRRs are predominantly expressed by innate immune cells, there is an emerging evidence that DAMP system is also active in fibroblasts (Bautista-Hernandez et al., 2017). For instance, ECM component tenascin-C can activate TLR4 signaling in synovial fibroblasts and induce pro-inflammatory cytokine production in the mouse model of rheumatoid arthritis (Midwood et al., 2009), while in the heart DAMP signaling activates an inflammatory and profibrotic response by cardiac fibroblasts (Turner, 2016; Zhang et al., 2015b). Fibroblasts also contribute to DAMP response by the surrounding innate immune cells and other tissue-resident cell types by promoting ECM degradation and fragmentation via secretion of ECM remodeling enzymes. Indeed, following proteolytic cleavage, many ECM fragments can bind to PRRs (McQuitty et al., 2020). For example, cleaved low molecular weight hyaluronan (LMW-HA), an abundant extracellular polysaccharide, can bind to TLR2/4 and promote NF-kB mediated proinflammatory cytokine production (Lee-Sayer et al., 2015). Interestingly, ECM-derived DAMPs can also mediate the immunosuppressive and pro-repair functions of immune cells that are linked to fibrotic remodeling (Frevert et al., 2018). For example, while LMW-HA promotes inflammation by agonizing TLR2 signaling, high molecular weight hyaluronan (HMW-HA) inhibits TLR2 signaling and can promote immunosuppressive T regulatory cell action (Bollyky et al., 2007). Thus, fibroblasts contribute to inflammation and tissue repair via activation and responding to DAMP signals.
Organ-specific fibroblasts
Despite sharing similar properties and responding to many of the same signal transduction pathways, tissue specific fibroblast functions and lineages exist to support the developmental, homeostatic and repair needs of specific organs. These emerge during embryonic development as multiple cell lineages converge to form fibroblasts that populate organs arising from all three somatic germ layers, such as ectodermally derived skin and mammary gland, mesodermally derived skeletal or heart muscles, and endodermally derived lung. The majority of fibroblasts in the body derive from the precursors of paraxial mesoderm and lateral plate mesoderm, while fibroblasts in the craniofacial structures originate from the neural crest mesenchyme (Herriges and Morrisey, 2014; Soriano, 1997) (Figure 5). In the heart, epicardial and endocardial epithelial cells generate fibroblasts through epithelial-to-mesenchymal transition (EMT) and endothelial-to-mesenchymal transition (EndMT), respectively (Gittenberger-de Groot et al., 1998) (Figure 5D).
Figure 5. Organ-specific fibroblast organization and lineage relationships.
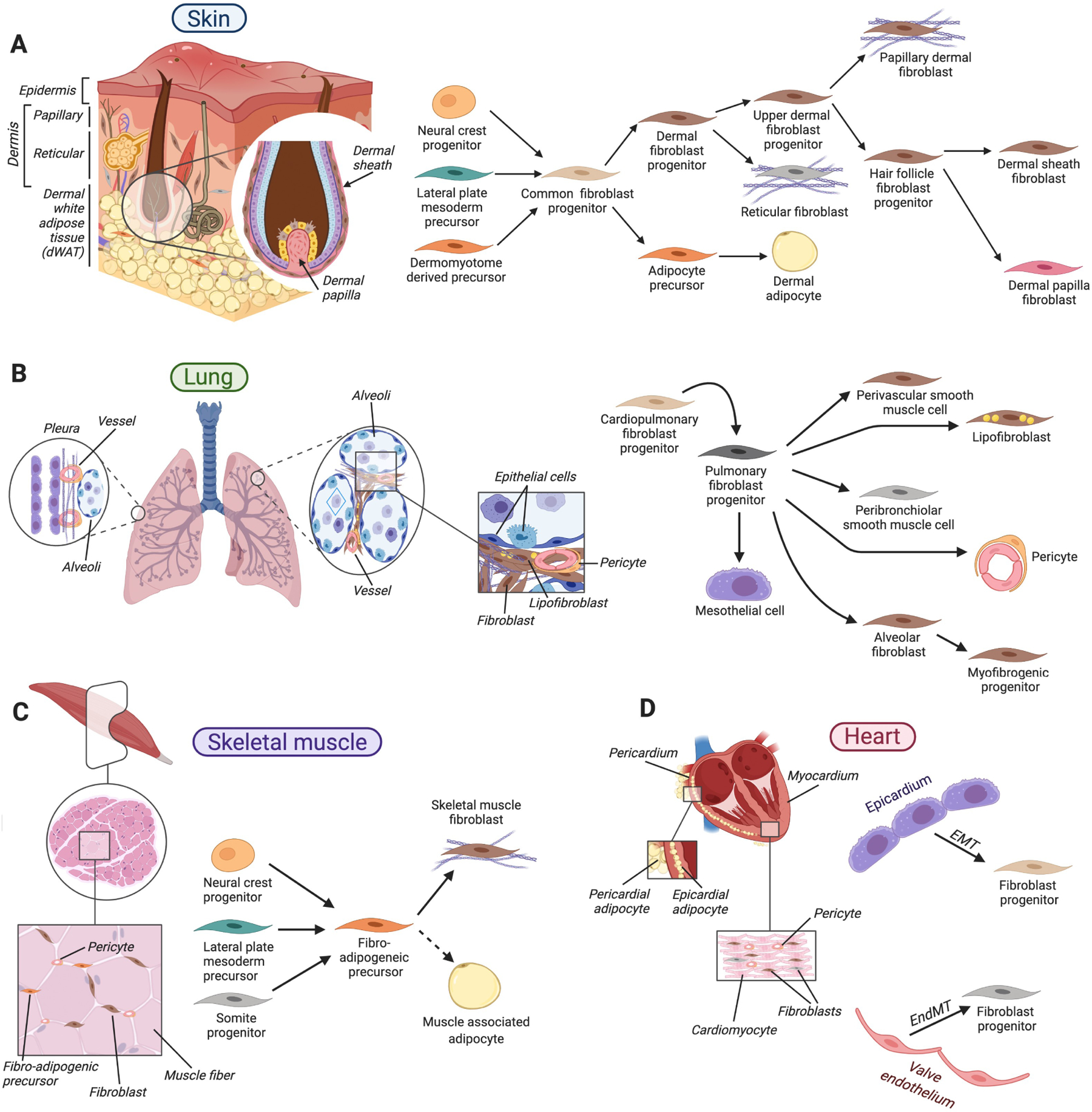
A. In the skin, diverse fibroblast types reside within its dermal layers, in dermal white adipose tissue and in association with hair follicles. During skin development, a common mesenchymal progenitor gives rise to dermal fibroblast progenitors that further specify toward fibroblasts of papillary and reticular dermis and hair follicle-associated fibroblasts, and to adipocyte precursors of dermal adipose tissue. B. In the lung, diverse fibroblasts associate with alveoli at the end of the branched epithelium, bronchioles and vasculature. During lung development, common cardiopulmonary progenitors generate a diverse array of fibroblasts, including mesothelial cells of the pleura, alveolar fibroblasts that support gas-exchanging epithelium, lipid droplet-containing lipofibroblasts, peribronchiolar and perivascular smooth muscle cells. C. Skeletal muscle fibroblasts, fibro-adipogenic precursors, and pericytes lie in the space between muscle fibers. Several developmentally distinct embryonic mesenchymal progenitors give rise to fibro-adipogenic precursors that, in turn, serve as long-lasting sources for muscle-associated fibroblasts as well as adipocytes upon aging and diseased states. D. Cardiac fibroblasts and pericytes reside between cardiomyocytes. During development, cardiac fibroblasts form from epicardial and endocardial epithelial cells via epithelial-to-mesenchymal transition (EMT) and endothelial-to-mesenchymal transition (EndMT), respectively. In A-D arrows indicate lineage relationships.
Once fibroblasts populate specific organs, they generate distinct microarchitecture, biophysical and biochemical components of connective tissues. Here, we will focus on four well-characterized organs with diverse architectural designs and physiology: two epithelium-rich organs – skin and lung, and two mesenchymal organs – skeletal muscle and heart. We will introduce the organization of these organs into their functional units, delineate fibroblast heterogeneity, and briefly discuss where fibroblasts are located within each unit and the molecular mechanisms that contribute to their development, function, and fibrosis.
Fibroblasts of the skin
Fibroblasts in the skin produce a mechanically resilient, adhesive yet elastic structural foundation which supports epithelial keratinocytes of the outward-facing stratified epidermis and its numerous appendages, primarily hair follicles and sweat glands. Fibroblasts and other mesenchymal lineages in the skin establish three anatomically distinct layers: papillary and reticular dermis and dermal white adipose tissue (dWAT) (Figure 5A). The epidermis is separated from the upper papillary dermis by a sheet of collagen and laminin-rich basement membrane jointly produced by interfacing epidermal keratinocytes and papillary fibroblasts. While in mice the papillary dermis in most body sites is very thin, in humans it forms complex undulating interdigitations with the epidermis, especially in the digit tips. Fibroblasts within often very thick reticular dermis produce densely packed ECM which endows skin with its mechanical strength. Fibroblast progenitors also produce and maintain dWAT, which contains a fine, web-like ECM that encases clusters of lipid-filled adipocytes. All three dermal layers also contain immune cells, epidermal appendages, sensory neurons, blood and lymphatic vessels, creating a complex network of cell communities that jointly support skin’s protective functions.
During embryonic development in mice, early mesenchymal precursors give rise to a common skin fibroblast progenitor, marked by the expression of the transcriptional regulators Engrailed 1 (En1) or Hypermethylated in cancer 1 (Hic1), or the transmembrane Protein delta homolog 1 (Dlk1) (Abbasi et al., 2020; Driskell et al., 2013; Rinkevich et al., 2015). After establishing a nascent connective tissue, skin fibroblast progenitors undergo progressive specification into distinct fibroblast lineages that assume discrete anatomical niches and express distinct molecular markers. Specifically, papillary fibroblasts in neonatal mouse skin express transmembrane factors CD26 (also known as Dpp4), Lrig1 and integrin Itga8, as well as the transcriptional repressor Prdm1 (also known as Blimp1). At the same time, precursors of reticular fibroblasts and dermal adipocytes express transcriptional factors Pparγ, Ebf2 as well as transmembrane proteins Pdpn and multifunctional signaling factor Sca1 (also known as Atxn1) (Driskell et al., 2013). As skin morphogenesis completes in early postnatal time and adult homeostasis ensues, fibroblasts undergo maturation and change their marker expression patterns (Driskell et al., 2013; Rognoni et al., 2016). For example, unlike in fetal skin, transcriptional regulator Lef1 becomes a gene marker of adult mouse papillary fibroblasts (Phan et al., 2020).
Despite papillary and reticular dermis being clearly demarcated from one another, mouse studies reveal the distribution of transcriptionally overlapping fibroblasts across skin layers (Shook et al., 2018). This observation suggests that distinct features and functions of dermal layers are the composite result of many cell types, and several fibroblast types working together. Recent scRNA-seq studies also rapidly increase our appreciation for fibroblast heterogeneity in the human skin. When carefully compared across several independent human skin scRNA-seq datasets, more than 90% of all dermal fibroblasts can be assigned to one of three shared groups, marked by the expression of WNT pathway members SFRP1 and SFRP2, as well as fat-binding Apolipoprotein E (APOE), respectively (Ascension et al., 2020). Each of these fibroblast groups can then be further subdivided, collectively resulting in as many as ten sub-groups with distinct marker gene combinations. Future work is needed to define the functional role of these heterogenous fibroblast populations.
Skin appendages, especially hair follicles, are populated by highly specialized fibroblasts whose major function is to regulate epithelial lineage activities, including stem cell quiescence, proliferation and differentiation. In the hair follicle, these include the so-called dermal papilla (DP) fibroblasts that form a tight cell cluster at its base, and the fibroblasts of the dermal sheath (DS) that encases hair follicle from the outside (Figure 5A). Both types of fibroblasts develop during hair follicle morphogenesis from embryonic papillary precursors via an intermediary dermal condensate progenitor (Mok et al., 2019; Sennett et al., 2015) and once fully formed, they express several specific marker genes (Driskell et al., 2011; Rezza et al., 2016; Shin et al., 2020) (Tables 1, 2). Compared to other skin fibroblasts, DP fibroblasts have several distinctive features. First, they are highly aggregative, including in vitro, where they form mound-like colonies before reaching confluence (Jahoda and Oliver, 1984). Second, they specifically associate with and function as the signaling niche for epithelial hair stem cells. Third, their gene expression prominently and periodically changes in synchrony with the hair growth cycle – a repetitive process of growing hairs that consists of dormant, active and regression phases. The resulting changes in DP fibroblast secretome critically drive transitions between cycle phases (Sennett and Rendl, 2012). Fourth, unlike other adult skin fibroblasts, DP fibroblasts can induce formation of new hair follicles when recombined with appropriately competent skin epithelium (Guerrero-Juarez et al., 2018; Reynolds and Jahoda, 1992). Lastly, just like hair follicles themselves, DP fibroblasts are highly heterogeneous across body regions and within the same region. For example, mouse DP fibroblasts that associate with different hair shapes (e.g. straight vs. zigzag) exhibit unique gene expression patterns (Driskell et al., 2009; Rezza et al., 2016). Analogous to mice, DP fibroblasts in humans play critical roles in hair follicle morphogenesis and cyclic growth (Higgins et al., 2013; Oh et al., 2016) and differences in the transcriptional response by DP fibroblasts to androgens across scalp skin regions underlie the pathogenesis of male and female pattern baldness (Chew et al., 2016).
Table 1:
Murine and Human Fibroblast Markers
| Rodent Skin Fibroblast Diversity | ||
|---|---|---|
| Fibroblast subsets | Markers | References |
| Pan-fibroblast | Pdgfrα+ Dpt+ Col1a2+ Col3a1+ Twist2+ Vimentin+ |
Collins et al. 2011; Driskell et al. 2013; Guerrero-Juarez et al. 2019; Abbasi et al. 2020; Phan et al. 2020 |
| Putative common dermal fibro-adipogenic progenitor | En1+ Dlk1+ Hic1+ |
Rinkevich et al. 2016; Abbasi et al. 2020; Driskell et al. 2013 |
| Papillary dermal fibroblast | CD26/Dpp4+ Dlk1neg Sca1/Ly6aneg Blimp1+ EphB2+ Lrig1+ Trps1+ |
Driskell et al. 2013; Wojciechowicz et al. 2013; Egawa et al. 2009 |
| Reticular dermal fibroblast | CD26/Dpp4neg Dlk1+ Sca1/Ly6aneg |
|
| Hypodermal fibroblast | CD26neg Dlk1neg Sca1/Ly6a+/− |
|
| Dermal sheath fibroblast | αSMA+ Itgα8+ Itgα5+ Col11a1+ Acan+ CD200+ Myh10+ Mlck+ Myl9+ |
Collins et al. 2011; Hagner et al. 2020; Heitman et al. 2020; Rahmani et al. 2014; Shin et al. 2020 |
| Dermal papilla fibroblast | Sox2+ Lef1+ Crabp1+ Rspo3+ Corin+ Versican+ Alkaline phosphatase+ |
Biernaskie et al. 2009; Driskell et al. 2009; Hagner et al. 2020; Sennet et al. 2015; Kishimoto et al. 1999; Rendl et al. 2005 |
| Hair follicle dermal stem cell (hfDSC) | Sox2+ αSMA+ Itgα8+ Mgp+ Acan+ |
Rahmani et al. 2014; Hagner et al. 2020; Shin et al. 2020 |
| Reversible fibroblast states | Marker status | References |
| Myofibroblast | αSMA+ Sm22+ ED-A fibronectin+ |
Plikus et al. 2017; Serini et al. 1998; Gabbiani et al. 1973 |
| Wound-induced regeneration-competent state | Crabp1+ Desmin+ Prss35+ |
Guerrero-Juarez et al. 2019; Abbasi et al. 2020; Phan et al. 2020; Lim et al. 2018 |
| Human Skin Fibroblast Diversity | ||
| Fibroblast subsets | Markers | References |
| Pan-fibroblast | COL1A1+ DCN+ CD90+ PDGFRα+ PDGFRβ+ LUM+ DCN+ MFAP5+ Vimentin+ Lineageneg (CD31neg CD45neg E-cadneg) |
Philippeos et al. 2018; Solé-Boldo et al. 2020; Vorstandlechner et al. 2020 |
| Papillary dermal fibroblast | COL6A5+ APCDD1+ HSPB3+ CD39+ CD26/DPP4+/− |
Philippeos et al. 2018; Solé-Boldo et al. 2020; Vorstandlechner et al. 2020; Tabib et al. 2018 |
| Reticular dermal fibroblast | CD36+ CTHRC1+ CD26/DPP4+/− |
|
| Dermal sheath fibroblast | ITGα5+ αSMA+ |
Hagner et al. 2020; Jahoda et al. 1991 |
| Dermal papilla fibroblast | RUNX1+ VCAN+ Androgen receptor+ 5-α-Reductase+ Alkaline phosphatase+ |
Hagner et al. 2020; Rutberg et al. 2006; Eicheler et al. 1998 |
| Reversible fibroblast states | Marker status | References |
| Myofibroblasts | αSMA+ ED-A fibronectin+ |
Bagalad et al. 2017 |
| Rodent Lung Fibroblast Diversity | ||
| Fibroblast subsets | Markers | References |
| Cardiopulmonary fibroblast progenitor | Wnt2+ Gli1+ Isl1+ |
Peng et al. 2013 |
| Pulmonary fibroblast progenitor | Tbx4+ | Arora et al. 2012; Xie et al. 2016; Zhang et al. 2013 |
| Alveolar fibroblast | Pdgfrα+ Col1a1+ Npnt+ Ces1d+ |
Lao et al. 2016; Tsukui et al. 2020; Xie et al. 2018 |
| Adventitial fibroblast | Pdgfrα+
Col1a1+ Pi16+ Ccl11+ |
|
| Peribronchial fibroblast | Pdgfrα+
Col1a1+ Hhip+ |
|
| Lipofibroblast | Vim+ Col4a1+ Fabp1+ Fabp4+ Lpl+ Fabp5+ Lipa+ Pparg+ Plin2+ |
Varisco et al. 2012; El Agha et al. 2017; Xie et al. 2018; Park et al. 2019; Chen et al. 2012; Imamura et al. 2002 |
| Myofibrogenic progenitor | Axin2+ | Zepp et al. 2018 |
| Rodent Skeletal Muscle Fibroblast Diversity | ||
| Fibroblast subsets | Markers | References |
| Fibro-adipogenic precursor (FAP) | Tcf4+
Gli1+ Sca1/Ly6a+ Pdgfrα+ Hic1+ |
Murphy et al. 2011; Scott et al. 2019; Joe et al. 2010; Kramann et al. 2015 |
| Rodent Heart Fibroblast Diversity | ||
| Fibroblast subsets | Markers | References |
| Epicardium-derived fibroblast progenitor | Hic1+ Gli1+ Pdgfrα+ Sca1/Ly6a+ |
Soliman et al. 2020; Deb & Ubil, 2014; Forte et al. 2020 |
| Valve endothelium-derived fibroblast progenitor | Wif1+ Dkk3+ |
|
| Rodent Pericyte Markers | ||
| Pericyte | Pdgfrβ+ Tbx18+ Ng2/CSPG4+ Glast1+ Rgs5+ Desmin+ αSMA+/− CD13/Anpep+ |
Lindahl et al. 1997; Guimarães-Camboa et al. 2017; Ozerdem et al. 2001; Zhu et al. 2008; Bondjers et al. 2003; Cho et al. 2003; Nehls et al. 1992; Dermietzel & Krause, 1991 |
Table 2:
Transgenic Mouse Lines Used to Mark Fibroblast Lineages in Skin, Heart, Lungs, and Skeletal Muscle
| Skin | ||
|---|---|---|
| Mouse line | Lineage and temporal specificity | Reference |
| Pdgfra-H2B | Pan-fibroblast | Collins et al. 2011 |
| Pdgfra-CreER | Pan-fibroblast | Driskell et al. 2013 |
| Blimp1-Cre | Arrector pili muscle /Papillary fibroblasts /Dermal papilla cells /Dermal sheath cells /Epidermis /Blood vessels | Driskell et al. 2013 |
| Dlk1-CreER | Reticular /Hypodermal fibroblasts (specific if labeled at E16.5-P2) | Driskell et al. 2013 |
| En1-Cre | Pan-fibroblast /Pericytes | Atit et al. 2006; Rinkevich et al. 2015 |
| Twist2/Dermo1-Cre | Pan-fibroblast | Atit et al. 2006; Phan et al. 2020 |
| Adipoq-Cre | dWAT /Adipose tissue | Rivera-Gonzalez et al. 2016; Schmidt & Horsley, |
| Lrig1-CreER | Papillary dermis /Epidermis (specific if labeled at E16.5) | Driskell et al. 2013 |
| Hic1-CreER | Pericytes /Dermal sheath cells /Papillary dermis /Reticular dermis /Hypodermis | Abbasi et al. 2020 |
| Col1a2-CreER | Pan-fibroblast | Zimmermann et al. 2014; Sun et al. 2020 |
| αSMA-CreERT2 | Dermal sheath cells /vascular smooth muscle cells (vSMCs) /Arrector pili muscle / myofibroblasts | Rahmani et al. 2014 |
| Tagln/Sm22-CreERT2 | Dermal sheath cells /vSMCs /Arrector pili muscle /myofibroblasts | Lim et al. 2018 |
| Prx1-Cre | Ventral dermal fibroblasts | Leavitt et al. 2020 |
| Sox2:GFP | Dermal papilla cells /Dermal sheath cells | Biernaskie et al. 2009; Driskell et al. 2009 |
| Acan-CreERT2 | Dermal sheath cells | Heitman et al. 2020 |
| Corin-CreER | Dermal papilla cells /Dermal sheath cells | Enshell-Seijffers et al. 2010 |
| Heart | ||
| Mouse line | Lineage and temporal specificity | Reference |
| Col1a2-CreER | Pan-fibroblast | Ubil et al. 2014; Pillai et al. 2017 |
| S100a4/Fsp1-CreER | Pan-fibroblast | Pillai et al. 2017; Song et al. 2012 |
| Pdgfra-CreER | Pan-fibroblast | Soliman et al. 2020 |
| Gli1-CreERT2 | Cardiopulmonary progenitors /Perivascular mesenchymal progenitors | Peng et al. 2013; Kramann et al. 2015 |
| Wnt2-CreERT2 | Cardiopulmonary progenitors | Peng et al. 2013 |
| Wt1-CreER and -CreERT2 | Epicardial-derived fibroblasts | Forte et al. 2020; Moore-Morris et al. 2014; Ali et al. 2014 |
| Tie1-CreER | Endothelium /endocardium-derived cardiac fibroblasts | Zeisberg et al. 2007 |
| Tek/Tie2-CreER | Endothelium /endocardium-derived cardiac fibroblasts | Moore-Morris et al. 2014; Ali et al. 2014 |
| Tbx18-CreER | Epicardial-derived cardiac fibroblasts | Ali et al. 2014 |
| Pax3-CreER | Neural crest-derived cardiac fibroblasts | Ali et al. 2014 |
| Postn-CreER | Fibroblasts following an insult (i.e., pressure overload, infarction) | Kaur et al. 2016; Takeda et al. 2009 |
| Lung | ||
| Mouse line | Lineage and temporal specificity | Reference |
| Pdgfra-CreER | Lung interstitial fibroblasts and lipofibroblasts | Ntokou et al. 2015; Li et al. 2018; Xie et al. 2018 |
| Col1a2-CreER | Pan-fibroblast | Kitamura et al. 2011; Hoyles et al. 2010 |
| S100a4/Fsp1-CreER | Pan-fibroblast | Li et al. 2011 |
| Fgf10-CreERT2 | Lipofibroblast progenitors | El Agha et al. 2014 |
| Foxd1-CreERT2 | Lung interstitial fibroblasts and pericyte progenitors | Hung et al. 2013 |
| Wt1-CreER and -CreERT2 | Injury-activated and interstitial fibroblasts, alveolar /vascular smooth muscle cells | Karki et al. 2014; Dixit et al. 2013; Que et al. 2008; Cano et al. 2013 |
| Tcf21-CreERT2 | Interstitial fibroblasts /lipofibroblasts /smooth muscle progenitors | Park et al. 2019 |
| Tbx2-Cre | Fibroblasts /Endothelial cells /Mesothelium /Smooth muscle cells | Wojahn et al. 2019 |
| Tek/Tie2-CreER | Endothelial-derived fibroblasts post-injury | Hashimoto et al. 2010 |
| αSMA-CreER | Myofibroblasts /vSMCs | Wu et al. 2007 |
| Fgf18-CreERT2 | Alveolar myofibroblasts /lipofibroblasts /Mesothelium | Hagan et al. 2019 |
| Tbx4-CreER | (Myo)fibroblasts and vascular smooth muscle progenitors /Rare endothelial cells | Xie et al. 2016 |
| Skeletal Muscle | ||
| Mouse line | Lineage and temporal specificity | Reference |
| Hic1-CreER | Skeletal muscle mesenchymal progenitors | Scott et al. 2019 |
| Adam12-CreER | Perivascular myofibroblast progenitors | Dulauroy et al. 2012 |
| Tcf4-CreER | Skeletal muscle fibroblasts | Mathew et al. 2011 |
| Prx1-CreER | Diaphragm muscle’s connective tissue fibroblasts | Merrell et al. 2015 |
Likewise, gene expression by DS fibroblasts is distinct and, prominently, includes contractile proteins (Table 1) (Heitman et al., 2020; Shin et al., 2020). The latter are important during hair growth termination, when smooth muscle-like contraction by DS helps the shrinking hair follicle to properly remodel to its dormant shape (Heitman et al., 2020). Near its base, DS harbors the so-called hair follicle dermal stem/progenitor cells that assure long-term replenishment of DP and DS with new fibroblasts (Rahmani et al., 2014; Shin et al., 2020). With advanced age, self-renewal capacity of dermal stem cells diminishes, leading to their exhaustion and permanent hair follicle atrophy. Moreover, DS fibroblasts directly interface with skin-resident immune cells and produce immune suppressive factors such as TGF ligands β1 and β2 and immunomodulatory molecules, such as Programmed death-ligand 1 (PDL1) and CD200, which may contribute to the immune privilege of hair follicles, a property that guards them against autoimmune reactions (Paus et al., 2003).
Underlying the reticular dermis, fibroblast progenitors with adipogenic potential give rise to lipid-laden adipocytes of the dWAT layer, which displays several distinctive properties compared to other white adipose depots in the body (Zwick et al., 2018) (Figure 5A). In addition to responding to changes in systemic nutrient availability, dWAT also prominently responds to local signals from cyclically growing hair follicles. During active hair growth phase, dWAT expands by hypertrophy of preexisting adipocytes and hyperplasia from dermal adipose stem cells (Festa et al., 2011; Zhang et al., 2016). Growth promoting effect of hair follicles on dWAT is mediated by signals, including BMP and Hedgehog ligands (Plikus et al., 2008; Zhang et al., 2016). Following hair growth regression, dWAT shrinks in part from loss of lipid and possibly through dedifferentiation of mature adipocytes. dWAT cells also exert several reciprocal effects on hair follicles. During rest phase, dWAT supports hair stem cell quiescence by secreting BMP ligands (Plikus et al., 2008), while at the onset of new hair growth, dWAT stimulates DP fibroblast activity via PDGF ligands (Festa et al., 2011). Beyond hair growth, dWAT progenitors actively sense bacteria and respond by rapidly differentiating and secreting the antimicrobial peptide cathelicidin (Zhang et al., 2019; Zhang et al., 2015a), while lipid content of mature dermal adipocytes can modulate skin ECM homeostasis (Zhang et al., 2021).
Skin fibrosis can occur in response to many triggers, including thermal burn, mechanical trauma, infection, radiation or surgery, or in association with systemic diseases, such as scleroderma and graft-versus-host disease, and it can result in hypertrophic scarring, keloid scarring and contractures that can impede mobility and skin reinnervation. Multiple populations of dermal fibroblasts, perivascular cells as well as dedifferentiated mature dermal adipocytes are the source of activated myofibroblasts and fibrotic ECM in injured or diseased skin (Dulauroy et al., 2012; Marangoni et al., 2015). Proliferative expansion of dermal fibroblasts is a common component of skin fibrosis, however, in certain contexts, such as upon bleomycin-induced fibrosis in mice, dermal fibroblast numbers, in fact, decrease (Shook et al., 2018). This suggests that fibrosis can occur because of exacerbated ECM-production of fewer preexisting fibroblasts. Moreover, increasing fibroblast proliferation within permissive wound environments can be associated with enhanced regeneration, further suggesting that fibroblast overproduction alone is not a key determinant of fibrosis (Abbasi et al., 2020). Both genetic mutations, as well as aberrant inflammatory signaling, can drive excessive skin fibrosis. For example, skin in patients with systemic sclerosis shows upregulated production of interleukin IL-13 (Li et al., 2017), while in mouse models skin fibrosis can be driven by interleukin IL-21 or by matrix metalloproteinase MMP12 (Taylor et al., 2018). Multiple types of immune cells have been implicated in skin fibrosis, prominently T cells and macrophages. Studies on immune and non-immune drivers of skin fibrosis, many of which are beyond the scope of this review, are informing the search for new anti-fibrotic therapies. For example, dermal fibrosis can be ameliorated in mouse models upon inhibition of myofibroblast-activating TGFβ and integrin signaling, such as with neutralizing antibodies, or upon stimulation of dermal adipogenesis with small-molecule PPARγ agonist rosiglitazone (Shi-wen et al., 2010).
Lung fibroblasts
While analogous to skin in being an epithelium-rich tissue, the lung is developmentally, architecturally, and functionally distinct which impacts the extent to which their fibroblast populations differ from one another. Skin epithelium is ectodermally derived, and its connective tissue forms distinct layers with voluminous ECM primed to provide mechanical resilience. In contrast, lung epithelium is of endodermal origin and forms a highly branched tree that terminates in expandable sacs, called alveoli. Gas exchange function of the lung critically relies both on the alveolar epithelium forming close physical connections with extensive capillary networks, and on repetitive and rapid ability of the lung to expand and contract. These functions necessitate the lung’s reticular ECM that is optimally viscoelastic and yet sufficiently strong to withstand changes in the air pressure and prevent alveolar rupture and potentially lethal barotrauma. Such ECM is produced by a diverse group of pulmonary mesenchymal cells, that in development originate from a subset of migrating cardiac mesoderm (Peng et al., 2013) (Figure 5B). As the lung matures, pulmonary mesenchymal progenitors assume distinct spatial positions and functions, including peribronchiolar and perivascular smooth muscle cells, pericytes, numerous populations of interstitial fibroblasts, pneumocyte-supporting lipofibroblasts, and mesothelial cells which line the visceral pleura (Figure 5B).
Cellular genealogy and disease contributions of these heterogeneous mesenchymal lung populations have been largely defined in mice on the basis of genetic lineage tracing and, more recently, with scRNA-seq. Early mesodermal progenitors that give rise to nearly all mesenchymal cells in an uninjured adult lung and that serve as the source for αSMA+ myofibroblasts following lung injury can be marked on the basis of expression of transcriptional factor Tbx4 (Arora et al., 2012). In fetal mice, Hedgehog pathway member Gli1 labels mesenchymal lung cells except lipofibroblasts (Li et al., 2015). Gli1 expression in these progenitors is functionally important as they depend on Hedgehog signaling for expansion (Kugler et al., 2017). Upon maturation Gli1+ progenitors increase expression of shared fibroblast marker PDGFRα (Rock et al., 2011). Additional heterogeneity is recognized within PDGFRα+ lung fibroblasts and that includes a WNT responsive subset, identified on the basis of Axin2 expression (Rock et al., 2011). Lipid droplet-containing lipofibroblasts are a distinct mesenchymal lung cell population required for proper development of epithelial alveolar type 2 cells, storage of vitamin A and production of surfactant (Al Alam et al., 2015). Intriguingly, it remains unclear if the major population of Gli1+/PDGFRα+ fibroblasts is the dominant source of lung ECM. It is likely that other lung cell populations partake in ECM synthesis, since the latter remains largely intact following Gli1+ cell pruning.
Upon injury, epithelial cells in the adult lung downregulate Hedgehog signaling, which activates proliferation of epithelium and normally quiescent mesenchyme (Peng et al., 2013). While epithelial stem cells repair airway and alveolar tissue, mesenchymal cells both repair and constrict the interstitial stroma. Mouse studies have identified PDGFRα+/Axin2+ fibroblasts that respond to injury by proliferating within the alveolar niche, and PDGFRα−/Axin2+ fibroblasts that largely contribute to forming anatomically distinct set of parabronchial myofibroblasts. Among other pathways, WNT signaling is an important driver of injury response by mouse pulmonary fibroblasts, especially by Axin2-expressing cells, and this signaling requirement is conserved in humans, for example during pathogenesis of pulmonary lymphangioleiomyomatosis, a rare disease in which fibroblasts and other cells form nodules that leads to progressive decline of pulmonary function (Obraztsova et al., 2020).
Many forms of pulmonary fibrosis involve the inexorable, progressive obliteration of parenchymal tissue which ultimately impairs gas exchange to cause respiratory failure and death. While no animal model accurately recapitulates all aspects of human lung fibrosis, bleomycin-induced lung injury can transiently increase the abundance of αSMA+ myofibroblasts and ECM in alveolar regions (Rock et al., 2011). Lineage tracing in this mouse model implicated numerous mesenchymal cells as the source for fibrotic myofibroblasts, including PDGFRα+ fibroblasts, lipofibroblasts, pericytes, and WT1+ mesothelial cells (El Agha et al., 2017; Hung et al., 2013). Such cooperative contribution of several tissue-resident mesenchymal cell types to fibrosis parallels analogous observations in other organs, including skin responses to wounding. Intriguingly, the bulk of the fibrotic tissue-producing lung fibroblasts can be marked with and are regulated by Tbx4 (Xie et al., 2016), a transcriptional factor that also marks early mesodermal lung progenitors during embryonic morphogenesis (Arora et al., 2012). This suggests cooption of developmental transcriptional regulators by adult fibroblasts for injury response. The targeting of these cells for the treatment of human disease remains an active area of investigation.
Skeletal muscle fibroblasts
Distinct from skin and lung, voluntary striated muscles lack epithelial structures and instead consists of parallel arrays of large, highly differentiated and multinucleated muscle fibers, whose primary function is force generation via contraction. Whereas some muscle fibers terminate at the tendon junctions, many of them end within the intramuscular connective tissue and transmit their contractile forces laterally. Intramuscular connective tissue has complex and hierarchical organization. Its innermost layer, called endomysium, surrounds individual muscle fiber and contains specialized laminin- and type IV collagen-rich basement membrane. Acting via its transmembrane receptors, laminin aids in mechanical force transduction from the intracellular contractile apparatus to the outer endomysium layer, rich in type I and III collagens (Chapman et al., 2016). Groups of myofibers, called fascicles, are surrounded by perimysium, which forms a continuum with tendons and is rich in the so-called perimysium “cables” – thick connective tissue bundles primarily composed of tightly packed type I and III collagen III fibrils. Finally, the entire muscle is enveloped by the epimysium connective layer (Chapman et al., 2016). The major role of perimysium in force transduction and its distinct ECM is supported by specialized perimysial fibroblasts that express high levels of thrombospondins 1 and 4, and type XI collagen. Perimysial fibroblasts express a number of matrisome and non-matrisome genes shared with tendon and cartilage and on scRNA-seq display substantial additional heterogeneity, whose functional significance remains to be understood (Muhl et al., 2020). A distinct type of perimysial fibroblasts, that expresses high levels of periostin, uniquely localizes at the perimysium-endomysium boundary and on scRNA-seq shares similarity with a constellation of endomysial mesenchymal cells, which include perivascular fibroblasts and fibro-adipogenic progenitors, that have a lineage potential to differentiate toward lipid-laden adipocytes (Figure 5C). Similar to skin and heart muscle, which we discuss below, skeletal muscle fibroblasts with progenitor properties express Sca1 (Joe et al., 2010; Uezumi et al., 2010), and similar to lung and heart – Gli1 (Kramann et al., 2015. Further, as with the heart, transcriptional repressor HIC1 has been identified as the regulator of quiescence for skeletal muscle fibroblasts (Scott et al., 2019).
Skeletal muscles are highly regenerative, a property that is supported by the so-called satellite stem cells, that are myogenic and express PAX7 (Relaix and Zammit, 2012). In response to muscle injury, both satellite stem cells and the surrounding interstitial fibroblasts activate and expand concurrently. Normally, satellite cells should expand more to give rise to new myoblasts that upon fusion produce new myofibers. However, upon the satellite cell deficiency that can be experimentally induced in mice through genetic means or normal aging, muscle fiber regeneration becomes deficient. In response, fibroblasts - including fibro-adipogenic progenitors - expand and generate excessive fibrotic ECM and excessive intramuscular adipocytes that become collectively detrimental to muscle’s contractile property. Conversely, if fibroblasts are depleted, premature myoblast differentiation, smaller myofibers, and rapid depletion of the satellite cell reserve occurs (Murphy et al., 2011). Therefore, fibroblasts do not merely compete for tissue space with satellite cells, but form a signaling niche, necessary for long-term maintenance and proper cellular dynamics of the myogenic lineage. Niche function of muscle fibroblasts parallels that played by DP fibroblasts in the context of hair follicle epithelial stem cells.
Notably, in response to injury, activated skeletal muscle fibroblasts undergo dynamic gene expression changes that are analogous to these in other organs, such as skin or heart. First, they express cytokines and other proinflammatory factors, followed by genes involved in the cell cycle and ECM. The latter enable activated myofibroblasts to rapidly deposit a provisional matrix, which then provides a new scaffold for regenerating myofibers. As muscle regenerates, the provisional matrix becomes rapidly remodeled and then almost completely removed, leaving space for the newly expanding myofibers (Joe et al., 2010; Scott et al., 2019). In this regard, regeneration of myofibers over the provisional ECM that rapidly remodels partially parallels the ability of large skin wounds to regenerate new hair follicles and adipocytes, both of which form within a provisional scar tissue.
Fibroblasts of the heart
Heart muscle is an anatomically and physiologically complex contractile organ, whose major cell population, cardiomyocytes connect with one another to form an electrically coupled tissue via intercalated disks that constitute its middle layer called myocardium. Heart is also rich in fibroblasts that generate and remodel a robust ECM network essential for electrical conductivity and heartbeat rhythm. In addition to myocardium, fibroblasts populate the outermost epicardium layer, which contains specialized adipose tissue, as well as the innermost endocardium, which is bordered by a layer of endothelial cells. In contrast to fibroblasts in most other organs that originate from mesenchymal progenitors via progressive specification, the majority of cardiac fibroblasts form via epithelial-to-mesenchymal transition (EMT) and those residing in the interventricular septum and right ventricle are the product of endothelial-to-mesenchymal transition (EndMT) (Gittenberger-de Groot et al., 1998) (Figure 5D).
As in other organs, adult cardiac fibroblasts are heterogeneous, and their lineage contributions are distinct. Single-cell transcriptomic studies on adult murine cardiac fibroblasts consistently reveal two main populations (Skelly et al., 2018) A smaller population of endocardial-derived fibroblasts expresses WNT signaling factors Wif1 and Dkk3 and presents a gene signature related to valve leaflets and, intriguingly, endochondral specification toward the bone lineage. A larger population of epicardial-derived fibroblasts further dichotomizes into groups characterized by either high expression of genes associated with metabolism, or genes associated with cell migration. The latter single-cell fibroblast population is likely identical to previously isolated fibroblast population that expressed markers PDGFRα and Sca1 and displays high clonogenic properties (Chong et al., 2011).
Considering that heart muscle is poorly regenerative, and that injury typically results in repair with a functionally deficient, non-contractile scar, much effort has focused on understanding cardiac fibroblasts in the context of fibrosis. In response to an acute injury, such as myocardial infarction, cardiac fibroblasts rapidly activate expression of chemoattractant, proinflammatory and profibrotic signals (Forte et al., 2020). Epicardial adipocytes in mice can also promote inflammation, such as via release of fatty acids (Chang et al., 2018). Similar to acute wounds in other organs, such as skin, murine cardiac fibroblasts in the infarction site become contractile (i.e. myofibroblasts) and transiently proliferative (Fu et al., 2018). They acquire an elongated aspect ratio and express distinct ECM proteins, including Cartilage oligomeric matrix protein (Comp) and thrombospondin 4, which are normally restricted to skeletal elements, such as bones, tendons or cartilages. These and other related ECM factors are thought to confer added strength to the mature cardiac scar, which is critical for its longevity within the constantly contracting heart. As cardiac scar remodels, fibroblast populations distal of the infarcted muscle also display changes consistent with compensatory interstitial fibrosis, including expression of ECM proteins and matrix remodeling factors.
When lineages are considered, injury to the myocardium activates both epicardial- and endocardial-derived fibroblast populations (Moore-Morris et al., 2014), yet the exact cellular origin of cardiac fibrosis has yet to be fully resolved. The essential role of PDGFRα+ fibroblasts in cardiac fibrosis is supported by mouse studies in which deletion of Hic1 in Pdgfra-expressing cells results in epicardium thickening, interstitial fibrosis and intra-myocardial adipogenesis in the absence of damage (Soliman et al., 2020). Other studies in mice have suggested that heterogeneous populations of PDGFRα+ fibroblasts are important for cardiac fibrosis development, including epicardial-derived Sca1+ Gli1+ (Farbehi et al., 2019; Soliman et al., 2020), as well as Fibroblast activation protein α-expressing subsets. Clinically relevant marker podoplanin (Pdpn) is highly expressed in human ischemic cardiomyopathy patients and when inhibited in mice, post-infarction repair is promoted (Cimini et al., 2019).
Fibroblast-targeting therapies
Promoting regenerative healing
While robust mechanisms exist to repair injuries in mammals, they often culminate with fibrosis, which presents a major clinical challenge. Currently approved antifibrotic therapies target fibroblast activation in established disease and are not able to restore the architecture and function of diseased tissues. Thus, the development of strategies aimed at promoting repair of injured tissues would significantly advance the field. In this light, chronic non-healing wounds of the skin offer an opportunity to intervene because they can be diagnosed at an early stage. To date, however, the only FDA approved growth factor for chronic diabetic foot ulcers is the gel preparation of low doses of recombinant PDGF, which in clinical studies significantly decreases the time to healing (Steed, 2006). Yet, this treatment has limited impact on other forms of non-healing skin ulcers, such as pressure ulcers (Yamakawa and Hayashida, 2019). Several other studies have shown promising results with other growth factors in animal models, but clinical trials did not prove effective in humans, which may reveal the difference in tissue repair strategies between human and murine skin (Yamakawa and Hayashida, 2019).
Mechanical tension may also be a therapeutic target to improve tissue repair and promote regeneration. Deletion of the mechanically sensitive focal adhesion kinase (FAK) in fibroblasts can reduce scar formation in mice. Interestingly, mechanical offloading can improve wound repair in pigs (Gurtner et al., 2011) and a study inhibiting the mechanically sensitive transcription factor Yes-associated protein (YAP) in mouse skin fibroblasts promotes scarless wound healing and tissue regeneration, including hair follicle regeneration (Mascharak et al., 2021). Yap inhibition has also been shown to abrogate liver, lung, and kidney fibrosis in animal models, suggesting that targeting mechanically sensitive pathways might shift scarring repair to a more regenerative healing process.
While the plasticity of fibroblasts can be problematic in the context of fibrotic diseases, it also presents an opportunity for regenerative interventions. This realization has driven recent efforts to restore tissue anatomy and function by reprogramming fibroblasts in vivo in situ via molecular strategies, that include direct reprogramming. In the context of skin wounds, the innate ability of fibroblasts to become supportive of hair follicle regeneration can be induced by transcriptional reprogramming. For example, temporally induced supra-physiological activation of Hedgehog signaling in wound fibroblasts in mice can potently enhance their ability to acquire DP fibroblast identity, which results in regeneration of large numbers of hair follicle-like structures both in large and in small wounds (Lim et al., 2018). Similar results have also been achieved by deleting the transcriptional repressor Hic1 in fibroblasts (Abbasi et al., 2020) or following fibroblast-specific overexpression of Lef1 (Phan et al., 2020). “Deeper” trans-lineage reprogramming of fibroblasts into keratinocytes in skin wounds in situ is possible upon viral transduction with keratinocyte lineage-associated transcription factors Dnp63a Grhl2, Tfap2a and Myc, producing fibroblast-derived epidermis and enhancing wound re-epithelialization in mice (Kurita et al., 2018).
Analogous proof-of-principle examples of therapeutic in vivo reprogramming of fibroblasts have been shown in the liver and heart. Viral delivery of transcription factors, such as Foxa3, Gata4, Hnf1a and Hnf4a, to myofibroblasts in liver reprograms them into hepatocyte-like cells and reduces signs of liver fibrosis in the mouse model (Rezvani et al., 2016; Song et al., 2016). Likewise, virally delivered Gata4, Mef2c and Tbx5 induces direct reprogramming of heart-resident fibroblasts into cardiomyocytes in vivo, resulting in reduced fibrosis and improved cardiac function in a mouse model of myocardial infarction (Miyamoto et al., 2018). The above examples clearly highlight the therapeutic potential of direct reprogramming of tissue-resident fibroblasts into “worker” cells and suggest it as a novel scar-replacement strategy. However, several important issues, including efficiency of reprogramming factor delivery using non-integrative vectors and high fibroblast specificity if not exclusivity, will need to be solved for this attractive approach to be deemed safe for clinical applications.
Conclusions and Perspectives
The last several decades have seen remarkable progress in the understanding of fibroblast biology across organs and conditions. The field has progressed from phenotypic studies of cultured cells performed more than a century ago to complex genetic and functional observations in vivo that have been facilitated by new methods and techniques. These advances have revealed unexpected similarities and unique characteristics of fibroblasts across diverse organs such as the skin, lung, heart, and skeletal muscle that are currently being leveraged for the treatment of human disease.
Despite this progress, many questions and opportunities remain. For example, ongoing efforts employing single-cell multi-omic and spatial genomic technologies will provide key insights into fibroblast heterogeneity within and across tissues as well as their ability to assume multiple functional states in response to physiological or disease triggers. As our appreciation of essential fibroblasts’ functions extends well beyond ECM synthesis, so does the excitement over novel therapeutic possibilities that modulating them in situ can bring for a broad spectrum of diseases characterized by aging, pathologic remodeling, and tissue fibrosis. Emerging areas of investigation, including cross-regulation between wound fibroblasts, immune cells, and peripheral nervous system will be critical for understanding how to restore tissues to their pre-injury state. Finally, effective clinical translation will require rigorous verification in native human tissues and in human-like models such as organotypic cultures, xenografts and pluripotent cell-derived organoids. These and other advances will enable new approaches to the prevention, treatment, and perhaps reversal of fibrotic conditions by the achievement of appropriate and controlled repair.
Acknowledgments
MVP is funded by NSF grant DMS-1951144, NIH grant U01-AR073159, LEO foundation grants LF-OC-20–000611 and LF-AW_RAM-19–400008; VH is funded by NIH, NIAMS grants R01-AR076938, R01-AR0695505, R01-AR075412. ELH is supported by NIH NHLBI awards R01-HL152677, R01-HL109233, and DOD PR170078P1 and the Greenfield Foundation. RRD is funded by NIH, NIAMS grant R01AR078743–01. JB is supported by CIHR grant PJT-159487 and the Calgary Firefighters Burn Treatment Society. SS is supported by CIHR-Vanier and Killam Scholarships. SMT is supported by the Barry M Goldwater Scholarship. Figures were created using BioRender.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Declaration of Interests
The authors declare no competing interests.
References
- Abbasi S, Sinha S, Labit E, Rosin NL, Yoon G, Rahmani W, Jaffer A, Sharma N, Hagner A, Shah P. et al. (2020). Distinct Regulatory Programs Control the Latent Regenerative Potential of Dermal Fibroblasts during Wound Healing. Cell Stem Cell 27, 396–412 e396. [DOI] [PubMed] [Google Scholar]
- Al Alam D, El Agha E, Sakurai R, Kheirollahi V, Moiseenko A, Danopoulos S, Shrestha A, Schmoldt C, Quantius J, Herold S. et al. (2015). Evidence for the involvement of fibroblast growth factor 10 in lipofibroblast formation during embryonic lung development. Development 142, 4139–4150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armulik A, Genove G, and Betsholtz C (2011). Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev Cell 21, 193–215. [DOI] [PubMed] [Google Scholar]
- Arora R, Metzger RJ, and Papaioannou VE (2012). Multiple roles and interactions of Tbx4 and Tbx5 in development of the respiratory system. PLoS Genet 8, e1002866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ascension AM, Fuertes-Alvarez S, Ibanez-Sole O, Izeta A, and Arauzo-Bravo MJ (2020). Human dermal fibroblast subpopulations are conserved across single-cell RNA sequencing studies. J Invest Dermatol. [DOI] [PubMed]
- Asli NS, Xaymardan M, Forte E, Waardenberg AJ, Cornwell J, Janbandhu V, Kesteven S, Chandrakanthan V, Malinowska H, Reinhard H. et al. (2018). PDGFRα signaling in cardiac fibroblasts modulates quiescence, metabolism and self-renewal, and promotes anatomical and functional repair. bioRxiv, 225979.
- Balaban NQ, Schwarz US, Riveline D, Goichberg P, Tzur G, Sabanay I, Mahalu D, Safran S, Bershadsky A, Addadi L. et al. (2001). Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat Cell Biol 3, 466–472. [DOI] [PubMed] [Google Scholar]
- Bautista-Hernandez LA, Gomez-Olivares JL, Buentello-Volante B, and Bautista-de Lucio VM (2017). Fibroblasts: The Unknown Sentinels Eliciting Immune Responses Against Microorganisms. Eur J Microbiol Immunol (Bp) 7, 151–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bechtel W, McGoohan S, Zeisberg EM, Muller GA, Kalbacher H, Salant DJ, Muller CA, Kalluri R, and Zeisberg M (2010). Methylation determines fibroblast activation and fibrogenesis in the kidney. Nat Med 16, 544–550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernard K, Logsdon NJ, Ravi S, Xie N, Persons BP, Rangarajan S, Zmijewski JW, Mitra K, Liu G, Darley-Usmar VM, et al. (2015). Metabolic Reprogramming Is Required for Myofibroblast Contractility and Differentiation. J Biol Chem 290, 25427–25438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biressi S, Miyabara EH, Gopinath SD, Carlig PM, and Rando TA (2014). A Wnt-TGFbeta2 axis induces a fibrogenic program in muscle stem cells from dystrophic mice. Sci Transl Med 6, 267ra176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bollyky PL, Lord JD, Masewicz SA, Evanko SP, Buckner JH, Wight TN, and Nepom GT (2007). Cutting edge: high molecular weight hyaluronan promotes the suppressive effects of CD4+CD25+ regulatory T cells. J Immunol 179, 744–747. [DOI] [PubMed] [Google Scholar]
- Buechler MB, Pradhan RN, Krishnamurty AT, Cox C, Calviello AK, Wang AW, Yang YA, Tam L, Caothien R, Roose-Girma M. et al. (2021). Cross-tissue organization of the fibroblast lineage. Nature. [DOI] [PubMed]
- Burrows MT, and Neymann CA (1917). STUDIES ON THE METABOLISM OF CELLS IN VITRO : I. THE TOXICITY OFalpha-AMINO-ACIDS FOR EMBRYONIC CHICKEN CELLS. J Exp Med 25, 93–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cajal SR (1896). Manual de anatomía patológica general : seguida de un resumen de microscopia aplicada a la histología y bacteriología patológicas, 2nd edn (Madrid: N. Moya; ). [Google Scholar]
- Candille SI, Van Raamsdonk CD, Chen C, Kuijper S, Chen-Tsai Y, Russ A, Meijlink F, and Barsh GS (2004). Dorsoventral patterning of the mouse coat by Tbx15. PLoS Biol 2, E3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cappato S, Gamberale R, Bocciardi R, and Brunelli S (2020). Genetic and Acquired Heterotopic Ossification: A Translational Tale of Mice and Men. Biomedicines 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrel A (1912). On the Permanent Life of Tissues Outside of the Organism. J Exp Med 15, 516–528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang HX, Zhao XJ, Zhu QL, Hou Q, and Li Y (2018). Removal of epicardial adipose tissue after myocardial infarction improves cardiac function. Herz 43, 258–264. [DOI] [PubMed] [Google Scholar]
- Chang HY, Chi JT, Dudoit S, Bondre C, van de Rijn M, Botstein D, and Brown PO (2002). Diversity, topographic differentiation, and positional memory in human fibroblasts. Proc Natl Acad Sci U S A 99, 12877–12882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chapman MA, Meza R, and Lieber RL (2016). Skeletal muscle fibroblasts in health and disease. Differentiation 92, 108–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chew EGY, Tan JHJ, Bahta AW, Ho BS, Liu X, Lim TC, Sia YY, Bigliardi PL, Heilmann S, Wan ACA, et al. (2016). Differential Expression between Human Dermal Papilla Cells from Balding and Non-Balding Scalps Reveals New Candidate Genes for Androgenetic Alopecia. J Invest Dermatol 136, 1559–1567. [DOI] [PubMed] [Google Scholar]
- Chong JJ, Chandrakanthan V, Xaymardan M, Asli NS, Li J, Ahmed I, Heffernan C, Menon MK, Scarlett CJ, Rashidianfar A. et al. (2011). Adult cardiac-resident MSC-like stem cells with a proepicardial origin. Cell Stem Cell 9, 527–540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cimini M, Garikipati VNS, de Lucia C, Cheng Z, Wang C, Truongcao MM, Lucchese AM, Roy R, Benedict C, Goukassian DA, et al. (2019). Podoplanin neutralization improves cardiac remodeling and function after acute myocardial infarction. JCI Insight 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crider BJ, Risinger GM Jr., Haaksma CJ, Howard EW, and Tomasek JJ (2011). Myocardin-related transcription factors A and B are key regulators of TGF-beta1-induced fibroblast to myofibroblast differentiation. J Invest Dermatol 131, 2378–2385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DasGupta R, and Fuchs E (1999). Multiple roles for activated LEF/TCF transcription complexes during hair follicle development and differentiation. Development 126, 4557–4568. [DOI] [PubMed] [Google Scholar]
- de Lima JE, Blavet C, Bonnin M-A, Hirsinger E, Comai G, Yvernogeau L, Bellenger L, Mella S, Nassari S, Robin C. et al. (2020). BMP signalling directs a fibroblast-to-myoblast conversion at the connective tissue/muscle interface to pattern limb muscles. bioRxiv, 2020.2007.2020.211342.
- Demaria M, Ohtani N, Youssef SA, Rodier F, Toussaint W, Mitchell JR, Laberge RM, Vijg J, Van Steeg H, Dolle ME, et al. (2014). An essential role for senescent cells in optimal wound healing through secretion of PDGF-AA. Dev Cell 31, 722–733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dempsey TM, Sangaralingham LR, Yao X, Sanghavi D, Shah ND, and Limper AH (2019). Clinical Effectiveness of Antifibrotic Medications for Idiopathic Pulmonary Fibrosis. Am J Respir Crit Care Med 200, 168–174. [DOI] [PubMed] [Google Scholar]
- Denton CP, Khan K, Hoyles RK, Shiwen X, Leoni P, Chen Y, Eastwood M, and Abraham DJ (2009). Inducible lineage-specific deletion of TbetaRII in fibroblasts defines a pivotal regulatory role during adult skin wound healing. J Invest Dermatol 129, 194–204. [DOI] [PubMed] [Google Scholar]
- Desmouliere A, Chaponnier C, and Gabbiani G (2005). Tissue repair, contraction, and the myofibroblast. Wound Repair Regen 13, 7–12. [DOI] [PubMed] [Google Scholar]
- Dhouailly D, and Sengel P (1975). [Feather- and hair-forming properties of dermal cells of glabrous skin from bird and mammals]. C R Acad Hebd Seances Acad Sci D 281, 1007–1010. [PubMed] [Google Scholar]
- Dobaczewski M, Bujak M, Li N, Gonzalez-Quesada C, Mendoza LH, Wang XF, and Frangogiannis NG (2010). Smad3 signaling critically regulates fibroblast phenotype and function in healing myocardial infarction. Circ Res 107, 418–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Driskell RR, Clavel C, Rendl M, and Watt FM (2011). Hair follicle dermal papilla cells at a glance. J Cell Sci 124, 1179–1182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Driskell RR, Giangreco A, Jensen KB, Mulder KW, and Watt FM (2009). Sox2-positive dermal papilla cells specify hair follicle type in mammalian epidermis. Development 136, 2815–2823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Driskell RR, Lichtenberger BM, Hoste E, Kretzschmar K, Simons BD, Charalambous M, Ferron SR, Herault Y, Pavlovic G, Ferguson-Smith AC, et al. (2013). Distinct fibroblast lineages determine dermal architecture in skin development and repair. Nature 504, 277–281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dulauroy S, Di Carlo SE, Langa F, Eberl G, and Peduto L (2012). Lineage tracing and genetic ablation of ADAM12(+) perivascular cells identify a major source of profibrotic cells during acute tissue injury. Nat Med 18, 1262–1270. [DOI] [PubMed] [Google Scholar]
- Duong TE, and Hagood JS (2018). Epigenetic Regulation of Myofibroblast Phenotypes in Fibrosis. Curr Pathobiol Rep 6, 79–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebeling AH (1913). The Permanent Life of Connective Tissue Outside of the Organism. J Exp Med 17, 273–285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El Agha E, Moiseenko A, Kheirollahi V, De Langhe S, Crnkovic S, Kwapiszewska G, Szibor M, Kosanovic D, Schwind F, Schermuly RT, et al. (2017). Two-Way Conversion between Lipogenic and Myogenic Fibroblastic Phenotypes Marks the Progression and Resolution of Lung Fibrosis. Cell Stem Cell 20, 571. [DOI] [PubMed] [Google Scholar]
- Farbehi N, Patrick R, Dorison A, Xaymardan M, Janbandhu V, Wystub-Lis K, Ho JW, Nordon RE, and Harvey RP (2019). Single-cell expression profiling reveals dynamic flux of cardiac stromal, vascular and immune cells in health and injury. Elife 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrer RA, Saalbach A, Grunwedel M, Lohmann N, Forstreuter I, Saupe S, Wandel E, Simon JC, and Franz S (2017). Dermal Fibroblasts Promote Alternative Macrophage Activation Improving Impaired Wound Healing. J Invest Dermatol 137, 941–950. [DOI] [PubMed] [Google Scholar]
- Festa E, Fretz J, Berry R, Schmidt B, Rodeheffer M, Horowitz M, and Horsley V (2011). Adipocyte lineage cells contribute to the skin stem cell niche to drive hair cycling. Cell 146, 761–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forte E, Skelly DA, Chen M, Daigle S, Morelli KA, Hon O, Philip VM, Costa MW, Rosenthal NA, and Furtado MB (2020). Dynamic Interstitial Cell Response during Myocardial Infarction Predicts Resilience to Rupture in Genetically Diverse Mice. Cell Rep 30, 3149–3163 e3146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank-Bertoncelj M, Trenkmann M, Klein K, Karouzakis E, Rehrauer H, Bratus A, Kolling C, Armaka M, Filer A, Michel BA, et al. (2017). Epigenetically-driven anatomical diversity of synovial fibroblasts guides joint-specific fibroblast functions. Nat Commun 8, 14852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frevert CW, Felgenhauer J, Wygrecka M, Nastase MV, and Schaefer L (2018). Danger-Associated Molecular Patterns Derived From the Extracellular Matrix Provide Temporal Control of Innate Immunity. J Histochem Cytochem 66, 213–227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman SL, Sheppard D, Duffield JS, and Violette S (2013). Therapy for fibrotic diseases: nearing the starting line. Sci Transl Med 5, 167sr161. [DOI] [PubMed] [Google Scholar]
- Fu X, Khalil H, Kanisicak O, Boyer JG, Vagnozzi RJ, Maliken BD, Sargent MA, Prasad V, Valiente-Alandi I, Blaxall BC, et al. (2018). Specialized fibroblast differentiated states underlie scar formation in the infarcted mouse heart. J Clin Invest 128, 2127–2143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallini R, Lindblom P, Bondjers C, Betsholtz C, and Andrae J (2016). PDGF-A and PDGF-B induces cardiac fibrosis in transgenic mice. Exp Cell Res 349, 282–290. [DOI] [PubMed] [Google Scholar]
- Gay D, Ghinatti G, Guerrero-Juarez CF, Ferrer RA, Ferri F, Lim CH, Murakami S, Gault N, Barroca V, Rombeau I. et al. (2020). Phagocytosis of Wnt inhibitor SFRP4 by late wound macrophages drives chronic Wnt activity for fibrotic skin healing. Sci Adv 6, eaay3704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ge Y, Gomez NC, Adam RC, Nikolova M, Yang H, Verma A, Lu CP, Polak L, Yuan S, Elemento O. et al. (2017). Stem Cell Lineage Infidelity Drives Wound Repair and Cancer. Cell 169, 636–650 e614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gittenberger-de Groot AC, Vrancken Peeters MP, Mentink MM, Gourdie RG, and Poelmann RE (1998). Epicardium-derived cells contribute a novel population to the myocardial wall and the atrioventricular cushions. Circ Res 82, 1043–1052. [DOI] [PubMed] [Google Scholar]
- Gonzalez R, Moffatt G, Hagner A, Sinha S, Shin W, Rahmani W, Chojnacki A, and Biernaskie J (2017). Platelet-derived growth factor signaling modulates adult hair follicle dermal stem cell maintenance and self-renewal. NPJ Regen Med 2, 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gospodarowicz D (1974). Localisation of a fibroblast growth factor and its effect alone and with hydrocortisone on 3T3 cell growth. Nature 249, 123–127. [DOI] [PubMed] [Google Scholar]
- Gouveia L, Betsholtz C, and Andrae J (2018). PDGF-A signaling is required for secondary alveolar septation and controls epithelial proliferation in the developing lung. Development 145. [DOI] [PubMed] [Google Scholar]
- Guerrero-Juarez CF, Astrowski AA, Murad R, Dang CT, Shatrova VO, Astrowskaja A, Lim CH, Ramos R, Wang X, Liu Y. et al. (2018). Wound Regeneration Deficit in Rats Correlates with Low Morphogenetic Potential and Distinct Transcriptome Profile of Epidermis. J Invest Dermatol 138, 1409–1419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guerrero-Juarez CF, Dedhia PH, Jin S, Ruiz-Vega R, Ma D, Liu Y, Yamaga K, Shestova O, Gay DL, Yang Z. et al. (2019). Single-cell analysis reveals fibroblast heterogeneity and myeloid-derived adipocyte progenitors in murine skin wounds. Nat Commun 10, 650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gurtner GC, Dauskardt RH, Wong VW, Bhatt KA, Wu K, Vial IN, Padois K, Korman JM, and Longaker MT (2011). Improving cutaneous scar formation by controlling the mechanical environment: large animal and phase I studies. Ann Surg 254, 217–225. [DOI] [PubMed] [Google Scholar]
- Hata A, and Chen YG (2016). TGF-beta Signaling from Receptors to Smads. Cold Spring Harb Perspect Biol 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heitman N, Sennett R, Mok KW, Saxena N, Srivastava D, Martino P, Grisanti L, Wang Z, Ma’ayan A, Rompolas P. et al. (2020). Dermal sheath contraction powers stem cell niche relocation during hair cycle regression. Science 367, 161–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herriges M, and Morrisey EE (2014). Lung development: orchestrating the generation and regeneration of a complex organ. Development 141, 502–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higgins CA, Chen JC, Cerise JE, Jahoda CA, and Christiano AM (2013). Microenvironmental reprogramming by three-dimensional culture enables dermal papilla cells to induce de novo human hair-follicle growth. Proc Natl Acad Sci U S A 110, 19679–19688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinz B (2007). Formation and function of the myofibroblast during tissue repair. J Invest Dermatol 127, 526–537. [DOI] [PubMed] [Google Scholar]
- Hinz B (2010). The myofibroblast: paradigm for a mechanically active cell. J Biomech 43, 146–155. [DOI] [PubMed] [Google Scholar]
- Hinz B, Celetta G, Tomasek JJ, Gabbiani G, and Chaponnier C (2001). Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol Biol Cell 12, 2730–2741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinz B, Phan SH, Thannickal VJ, Prunotto M, Desmouliere A, Varga J, De Wever O, Mareel M, and Gabbiani G (2012). Recent developments in myofibroblast biology: paradigms for connective tissue remodeling. Am J Pathol 180, 1340–1355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hogue MJ (1919). The Effect of Hypotonic and Hypertonic Solutions on Fibroblasts of the Embryonic Chick Heart in Vitro. J Exp Med 30, 617–648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang X, Yang N, Fiore VF, Barker TH, Sun Y, Morris SW, Ding Q, Thannickal VJ, and Zhou Y (2012). Matrix stiffness-induced myofibroblast differentiation is mediated by intrinsic mechanotransduction. Am J Respir Cell Mol Biol 47, 340–348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hung C, Linn G, Chow YH, Kobayashi A, Mittelsteadt K, Altemeier WA, Gharib SA, Schnapp LM, and Duffield JS (2013). Role of lung pericytes and resident fibroblasts in the pathogenesis of pulmonary fibrosis. Am J Respir Crit Care Med 188, 820–830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hynes RO, and Naba A (2012). Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol 4, a004903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito M, Liu Y, Yang Z, Nguyen J, Liang F, Morris RJ, and Cotsarelis G (2005). Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat Med 11, 1351–1354. [DOI] [PubMed] [Google Scholar]
- Jahoda CA, and Oliver RF (1984). Vibrissa dermal papilla cell aggregative behaviour in vivo and in vitro. J Embryol Exp Morphol 79, 211–224. [PubMed] [Google Scholar]
- Joe AW, Yi L, Natarajan A, Le Grand F, So L, Wang J, Rudnicki MA, and Rossi FM (2010). Muscle injury activates resident fibro/adipogenic progenitors that facilitate myogenesis. Nat Cell Biol 12, 153–163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jorgenson AJ, Choi KM, Sicard D, Smith KM, Hiemer SE, Varelas X, and Tschumperlin DJ (2017). TAZ activation drives fibroblast spheroid growth, expression of profibrotic paracrine signals, and context-dependent ECM gene expression. Am J Physiol Cell Physiol 312, C277–C285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Junker JP, Sommar P, Skog M, Johnson H, and Kratz G (2010). Adipogenic, chondrogenic and osteogenic differentiation of clonally derived human dermal fibroblasts. Cells Tissues Organs 191, 105–118. [DOI] [PubMed] [Google Scholar]
- Kazlauskas A (2017). PDGFs and their receptors. Gene 614, 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruglikov IL, and Scherer PE (2017). Adipocyte-myofibroblast transition as a possible pathophysiological step in androgenetic alopecia. Exp Dermatol 26, 522–523. [DOI] [PubMed] [Google Scholar]
- Kugler MC, Loomis CA, Zhao Z, Cushman JC, Liu L, and Munger JS (2017). Sonic Hedgehog Signaling Regulates Myofibroblast Function during Alveolar Septum Formation in Murine Postnatal Lung. Am J Respir Cell Mol Biol 57, 280–293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuppe C, Ibrahim MM, Kranz J, Zhang X, Ziegler S, Perales-Paton J, Jansen J, Reimer KC, Smith JR, Dobie R. et al. (2020). Decoding myofibroblast origins in human kidney fibrosis. Nature. [DOI] [PMC free article] [PubMed]
- Kurita M, Araoka T, Hishida T, O’Keefe DD, Takahashi Y, Sakamoto A, Sakurai M, Suzuki K, Wu J, Yamamoto M. et al. (2018). In vivo reprogramming of wound-resident cells generates skin epithelial tissue. Nature 561, 243–247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lakos G, Takagawa S, Chen SJ, Ferreira AM, Han G, Masuda K, Wang XJ, DiPietro LA, and Varga J (2004). Targeted disruption of TGF-beta/Smad3 signaling modulates skin fibrosis in a mouse model of scleroderma. Am J Pathol 165, 203–217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le HQ, Ghatak S, Yeung CY, Tellkamp F, Gunschmann C, Dieterich C, Yeroslaviz A, Habermann B, Pombo A, Niessen CM, et al. (2016). Mechanical regulation of transcription controls Polycomb-mediated gene silencing during lineage commitment. Nat Cell Biol 18, 864–875. [DOI] [PubMed] [Google Scholar]
- Lee-Sayer SS, Dong Y, Arif AA, Olsson M, Brown KL, and Johnson P (2015). The where, when, how, and why of hyaluronan binding by immune cells. Front Immunol 6, 150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemos DR, and Duffield JS (2018). Tissue-resident mesenchymal stromal cells: Implications for tissue-specific antifibrotic therapies. Sci Transl Med 10. [DOI] [PubMed] [Google Scholar]
- Li C, Li M, Li S, Xing Y, Yang CY, Li A, Borok Z, De Langhe S, and Minoo P (2015). Progenitors of secondary crest myofibroblasts are developmentally committed in early lung mesoderm. Stem Cells 33, 999–1012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li G, Larregina AT, Domsic RT, Stolz DB, Medsger TA Jr., Lafyatis R, and Fuschiotti P (2017). Skin-Resident Effector Memory CD8(+)CD28(−) T Cells Exhibit a Profibrotic Phenotype in Patients with Systemic Sclerosis. J Invest Dermatol 137, 1042–1050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim CH, Sun Q, Ratti K, Lee SH, Zheng Y, Takeo M, Lee W, Rabbani P, Plikus MV, Cain JE, et al. (2018). Hedgehog stimulates hair follicle neogenesis by creating inductive dermis during murine skin wound healing. Nat Commun 9, 4903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lodyga M, and Hinz B (2020). TGF-beta1 - A truly transforming growth factor in fibrosis and immunity. Semin Cell Dev Biol 101, 123–139. [DOI] [PubMed] [Google Scholar]
- Lu P, Takai K, Weaver VM, and Werb Z (2011). Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb Perspect Biol 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacDonald BT, Tamai K, and He X (2009). Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell 17, 9–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marangoni RG, Korman BD, Wei J, Wood TA, Graham LV, Whitfield ML, Scherer PE, Tourtellotte WG, and Varga J (2015). Myofibroblasts in murine cutaneous fibrosis originate from adiponectin-positive intradermal progenitors. Arthritis Rheumatol 67, 1062–1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mascharak S, desJardins-Park HE, Davitt MF, Griffin M, Borrelli MR, Moore AL, Chen K, Duoto B, Chinta M, Foster DS, et al. (2021). Preventing Engrailed-1 activation in fibroblasts yields wound regeneration without scarring. Science 372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massague J (2012). TGFbeta signalling in context. Nat Rev Mol Cell Biol 13, 616–630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mastrogiannaki M, Lichtenberger BM, Reimer A, Collins CA, Driskell RR, and Watt FM (2016). beta-Catenin Stabilization in Skin Fibroblasts Causes Fibrotic Lesions by Preventing Adipocyte Differentiation of the Reticular Dermis. J Invest Dermatol 136, 1130–1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGee HM, Schmidt BA, Booth CJ, Yancopoulos GD, Valenzuela DM, Murphy AJ, Stevens S, Flavell RA, and Horsley V (2013). IL-22 promotes fibroblast-mediated wound repair in the skin. J Invest Dermatol 133, 1321–1329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McQuitty CE, Williams R, Chokshi S, and Urbani L (2020). Immunomodulatory Role of the Extracellular Matrix Within the Liver Disease Microenvironment. Front Immunol 11, 574276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meli VS, Atcha H, Veerasubramanian PK, Nagalla RR, Luu TU, Chen EY, Guerrero-Juarez CF, Yamaga K, Pandori W, Hsieh JY, et al. (2020). YAP-mediated mechanotransduction tunes the macrophage inflammatory response. Sci Adv 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Midwood K, Sacre S, Piccinini AM, Inglis J, Trebaul A, Chan E, Drexler S, Sofat N, Kashiwagi M, Orend G. et al. (2009). Tenascin-C is an endogenous activator of Toll-like receptor 4 that is essential for maintaining inflammation in arthritic joint disease. Nat Med 15, 774–780. [DOI] [PubMed] [Google Scholar]
- Millar SE (2002). Molecular mechanisms regulating hair follicle development. J Invest Dermatol 118, 216–225. [DOI] [PubMed] [Google Scholar]
- Miyamoto K, Akiyama M, Tamura F, Isomi M, Yamakawa H, Sadahiro T, Muraoka N, Kojima H, Haginiwa S, Kurotsu S. et al. (2018). Direct In Vivo Reprogramming with Sendai Virus Vectors Improves Cardiac Function after Myocardial Infarction. Cell Stem Cell 22, 91–103 e105. [DOI] [PubMed] [Google Scholar]
- Mok KW, Saxena N, Heitman N, Grisanti L, Srivastava D, Muraro MJ, Jacob T, Sennett R, Wang Z, Su Y. et al. (2019). Dermal Condensate Niche Fate Specification Occurs Prior to Formation and Is Placode Progenitor Dependent. Dev Cell 48, 32–48 e35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore-Morris T, Guimaraes-Camboa N, Banerjee I, Zambon AC, Kisseleva T, Velayoudon A, Stallcup WB, Gu Y, Dalton ND, Cedenilla M. et al. (2014). Resident fibroblast lineages mediate pressure overload-induced cardiac fibrosis. J Clin Invest 124, 2921–2934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison SJ, and Spradling AC (2008). Stem cells and niches: mechanisms that promote stem cell maintenance throughout life. Cell 132, 598–611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muhl L, Genove G, Leptidis S, Liu J, He L, Mocci G, Sun Y, Gustafsson S, Buyandelger B, Chivukula IV, et al. (2020). Single-cell analysis uncovers fibroblast heterogeneity and criteria for fibroblast and mural cell identification and discrimination. Nat Commun 11, 3953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy MM, Lawson JA, Mathew SJ, Hutcheson DA, and Kardon G (2011). Satellite cells, connective tissue fibroblasts and their interactions are crucial for muscle regeneration. Development 138, 3625–3637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naik S, Larsen SB, Gomez NC, Alaverdyan K, Sendoel A, Yuan S, Polak L, Kulukian A, Chai S, and Fuchs E (2017). Inflammatory memory sensitizes skin epithelial stem cells to tissue damage. Nature 550, 475–480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakajima H, Nakajima HO, Salcher O, Dittie AS, Dembowsky K, Jing S, and Field LJ (2000). Atrial but not ventricular fibrosis in mice expressing a mutant transforming growth factor-beta(1) transgene in the heart. Circ Res 86, 571–579. [DOI] [PubMed] [Google Scholar]
- Nakaya M, Watari K, Tajima M, Nakaya T, Matsuda S, Ohara H, Nishihara H, Yamaguchi H, Hashimoto A, Nishida M. et al. (2017). Cardiac myofibroblast engulfment of dead cells facilitates recovery after myocardial infarction. J Clin Invest 127, 383–401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obraztsova K, Basil MC, Rue R, Sivakumar A, Lin SM, Mukhitov AR, Gritsiuta AI, Evans JF, Kopp M, Katzen J. et al. (2020). mTORC1 activation in lung mesenchyme drives sex- and age-dependent pulmonary structure and function decline. Nat Commun 11, 5640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh JW, Kloepper J, Langan EA, Kim Y, Yeo J, Kim MJ, Hsi TC, Rose C, Yoon GS, Lee SJ, et al. (2016). A Guide to Studying Human Hair Follicle Cycling In Vivo. J Invest Dermatol 136, 34–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olson LE, and Soriano P (2009). Increased PDGFRalpha activation disrupts connective tissue development and drives systemic fibrosis. Dev Cell 16, 303–313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Opalenik SR, and Davidson JM (2005). Fibroblast differentiation of bone marrow-derived cells during wound repair. FASEB J 19, 1561–1563. [DOI] [PubMed] [Google Scholar]
- Pakshir P, Alizadehgiashi M, Wong B, Coelho NM, Chen X, Gong Z, Shenoy VB, McCulloch CA, and Hinz B (2019). Dynamic fibroblast contractions attract remote macrophages in fibrillar collagen matrix. Nat Commun 10, 1850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pangas SA, and Woodruff TK (2000). Activin signal transduction pathways. Trends Endocrinol Metab 11, 309–314. [DOI] [PubMed] [Google Scholar]
- Paus R, Ito N, Takigawa M, and Ito T (2003). The hair follicle and immune privilege. J Investig Dermatol Symp Proc 8, 188–194. [DOI] [PubMed] [Google Scholar]
- Peng T, Tian Y, Boogerd CJ, Lu MM, Kadzik RS, Stewart KM, Evans SM, and Morrisey EE (2013). Coordination of heart and lung co-development by a multipotent cardiopulmonary progenitor. Nature 500, 589–592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phan QM, Fine GM, Salz L, Herrera GG, Wildman B, Driskell IM, and Driskell RR (2020). Lef1 expression in fibroblasts maintains developmental potential in adult skin to regenerate wounds. Elife 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phan QM, Sinha S, Biernaskie J, and Driskell RR (2021). Single-cell transcriptomic analysis of small and large wounds reveals the distinct spatial organization of regenerative fibroblasts. Exp Dermatol 30, 92–101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pittenger MF, Discher DE, Peault BM, Phinney DG, Hare JM, and Caplan AI (2019). Mesenchymal stem cell perspective: cell biology to clinical progress. NPJ Regen Med 4, 22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, and Marshak DR (1999). Multilineage potential of adult human mesenchymal stem cells. Science 284, 143–147. [DOI] [PubMed] [Google Scholar]
- Plikus MV, Guerrero-Juarez CF, Ito M, Li YR, Dedhia PH, Zheng Y, Shao M, Gay DL, Ramos R, Hsi TC, et al. (2017). Regeneration of fat cells from myofibroblasts during wound healing. Science 355, 748–752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plikus MV, Mayer JA, de la Cruz D, Baker RE, Maini PK, Maxson R, and Chuong CM (2008). Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration. Nature 451, 340–344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahmani W, Abbasi S, Hagner A, Raharjo E, Kumar R, Hotta A, Magness S, Metzger D, and Biernaskie J (2014). Hair follicle dermal stem cells regenerate the dermal sheath, repopulate the dermal papilla, and modulate hair type. Dev Cell 31, 543–558. [DOI] [PubMed] [Google Scholar]
- Relaix F, and Zammit PS (2012). Satellite cells are essential for skeletal muscle regeneration: the cell on the edge returns centre stage. Development 139, 2845–2856. [DOI] [PubMed] [Google Scholar]
- Rendl M, Polak L, and Fuchs E (2008). BMP signaling in dermal papilla cells is required for their hair follicle-inductive properties. Genes Dev 22, 543–557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds AJ, and Jahoda CA (1992). Cultured dermal papilla cells induce follicle formation and hair growth by transdifferentiation of an adult epidermis. Development 115, 587–593. [DOI] [PubMed] [Google Scholar]
- Rezvani M, Espanol-Suner R, Malato Y, Dumont L, Grimm AA, Kienle E, Bindman JG, Wiedtke E, Hsu BY, Naqvi SJ, et al. (2016). In Vivo Hepatic Reprogramming of Myofibroblasts with AAV Vectors as a Therapeutic Strategy for Liver Fibrosis. Cell Stem Cell 18, 809–816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rezza A, Wang Z, Sennett R, Qiao W, Wang D, Heitman N, Mok KW, Clavel C, Yi R, Zandstra P. et al. (2016). Signaling Networks among Stem Cell Precursors, Transit-Amplifying Progenitors, and their Niche in Developing Hair Follicles. Cell Rep 14, 3001–3018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rinkevich Y, Walmsley GG, Hu MS, Maan ZN, Newman AM, Drukker M, Januszyk M, Krampitz GW, Gurtner GC, Lorenz HP, et al. (2015). Skin fibrosis. Identification and isolation of a dermal lineage with intrinsic fibrogenic potential. Science 348, aaa2151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rinn JL, Bondre C, Gladstone HB, Brown PO, and Chang HY (2006). Anatomic demarcation by positional variation in fibroblast gene expression programs. PLoS Genet 2, e119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivera-Gonzalez GC, Shook BA, Andrae J, Holtrup B, Bollag K, Betsholtz C, Rodeheffer MS, and Horsley V (2016). Skin Adipocyte Stem Cell Self-Renewal Is Regulated by a PDGFA/AKT-Signaling Axis. Cell Stem Cell 19, 738–751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rock JR, Barkauskas CE, Cronce MJ, Xue Y, Harris JR, Liang J, Noble PW, and Hogan BL (2011). Multiple stromal populations contribute to pulmonary fibrosis without evidence for epithelial to mesenchymal transition. Proc Natl Acad Sci U S A 108, E1475–1483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rognoni E, Gomez C, Pisco AO, Rawlins EL, Simons BD, Watt FM, and Driskell RR (2016). Inhibition of beta-catenin signalling in dermal fibroblasts enhances hair follicle regeneration during wound healing. Development 143, 2522–2535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sahai E, Astsaturov I, Cukierman E, DeNardo DG, Egeblad M, Evans RM, Fearon D, Greten FR, Hingorani SR, Hunter T. et al. (2020). A framework for advancing our understanding of cancer-associated fibroblasts. Nat Rev Cancer 20, 174–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sava P, Ramanathan A, Dobronyi A, Peng X, Sun H, Ledesma-Mendoza A, Herzog EL, and Gonzalez AL (2017). Human pericytes adopt myofibroblast properties in the microenvironment of the IPF lung. JCI Insight 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaefer L (2014). Complexity of danger: the diverse nature of damage-associated molecular patterns. J Biol Chem 289, 35237–35245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott RW, Arostegui M, Schweitzer R, Rossi FMV, and Underhill TM (2019). Hic1 Defines Quiescent Mesenchymal Progenitor Subpopulations with Distinct Functions and Fates in Skeletal Muscle Regeneration. Cell Stem Cell 25, 797–813 e799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sennett R, and Rendl M (2012). Mesenchymal-epithelial interactions during hair follicle morphogenesis and cycling. Semin Cell Dev Biol 23, 917–927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sennett R, Wang Z, Rezza A, Grisanti L, Roitershtein N, Sicchio C, Mok KW, Heitman NJ, Clavel C, Ma’ayan A. et al. (2015). An Integrated Transcriptome Atlas of Embryonic Hair Follicle Progenitors, Their Niche, and the Developing Skin. Dev Cell 34, 577–591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi M, Zhu J, Wang R, Chen X, Mi L, Walz T, and Springer TA (2011). Latent TGF-beta structure and activation. Nature 474, 343–349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi-wen X, Eastwood M, Stratton RJ, Denton CP, Leask A, and Abraham DJ (2010). Rosiglitazone alleviates the persistent fibrotic phenotype of lesional skin scleroderma fibroblasts. Rheumatology (Oxford) 49, 259–263. [DOI] [PubMed] [Google Scholar]
- Shi-wen X, Parapuram SK, Pala D, Chen Y, Carter DE, Eastwood M, Denton CP, Abraham DJ, and Leask A (2009). Requirement of transforming growth factor beta-activated kinase 1 for transforming growth factor beta-induced alpha-smooth muscle actin expression and extracellular matrix contraction in fibroblasts. Arthritis Rheum 60, 234–241. [DOI] [PubMed] [Google Scholar]
- Shin W, Rosin NL, Sparks H, Sinha S, Rahmani W, Sharma N, Workentine M, Abbasi S, Labit E, Stratton JA, et al. (2020). Dysfunction of Hair Follicle Mesenchymal Progenitors Contributes to Age-Associated Hair Loss. Dev Cell 53, 185–198 e187. [DOI] [PubMed] [Google Scholar]
- Shook BA, Wasko RR, Mano O, Rutenberg-Schoenberg M, Rudolph MC, Zirak B, Rivera-Gonzalez GC, Lopez-Giraldez F, Zarini S, Rezza A. et al. (2020). Dermal Adipocyte Lipolysis and Myofibroblast Conversion Are Required for Efficient Skin Repair. Cell Stem Cell 26, 880–895 e886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shook BA, Wasko RR, Rivera-Gonzalez GC, Salazar-Gatzimas E, Lopez-Giraldez F, Dash BC, Munoz-Rojas AR, Aultman KD, Zwick RK, Lei V. et al. (2018). Myofibroblast proliferation and heterogeneity are supported by macrophages during skin repair. Science 362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinha M, Sen CK, Singh K, Das A, Ghatak S, Rhea B, Blackstone B, Powell HM, Khanna S, and Roy S (2018). Direct conversion of injury-site myeloid cells to fibroblast-like cells of granulation tissue. Nat Commun 9, 936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skelly DA, Squiers GT, McLellan MA, Bolisetty MT, Robson P, Rosenthal NA, and Pinto AR (2018). Single-Cell Transcriptional Profiling Reveals Cellular Diversity and Intercommunication in the Mouse Heart. Cell Rep 22, 600–610. [DOI] [PubMed] [Google Scholar]
- Smith CL, Baek ST, Sung CY, and Tallquist MD (2011). Epicardial-derived cell epithelial-to-mesenchymal transition and fate specification require PDGF receptor signaling. Circ Res 108, e15–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soliman H, Paylor B, Scott RW, Lemos DR, Chang C, Arostegui M, Low M, Lee C, Fiore D, Braghetta P. et al. (2020). Pathogenic Potential of Hic1-Expressing Cardiac Stromal Progenitors. Cell Stem Cell 26, 205–220 e208. [DOI] [PubMed] [Google Scholar]
- Song G, Pacher M, Balakrishnan A, Yuan Q, Tsay HC, Yang D, Reetz J, Brandes S, Dai Z, Putzer BM, et al. (2016). Direct Reprogramming of Hepatic Myofibroblasts into Hepatocytes In Vivo Attenuates Liver Fibrosis. Cell Stem Cell 18, 797–808. [DOI] [PubMed] [Google Scholar]
- Sonnylal S, Denton CP, Zheng B, Keene DR, He R, Adams HP, Vanpelt CS, Geng YJ, Deng JM, Behringer RR, et al. (2007). Postnatal induction of transforming growth factor beta signaling in fibroblasts of mice recapitulates clinical, histologic, and biochemical features of scleroderma. Arthritis Rheum 56, 334–344. [DOI] [PubMed] [Google Scholar]
- Soriano P (1997). The PDGF alpha receptor is required for neural crest cell development and for normal patterning of the somites. Development 124, 2691–2700. [DOI] [PubMed] [Google Scholar]
- Steed DL (2006). Clinical evaluation of recombinant human platelet-derived growth factor for the treatment of lower extremity ulcers. Plast Reconstr Surg 117, 143S–149S; discussion 150S-151S. [DOI] [PubMed] [Google Scholar]
- Taylor DK, Mittereder N, Kuta E, Delaney T, Burwell T, Dacosta K, Zhao W, Cheng LI, Brown C, Boutrin A. et al. (2018). T follicular helper-like cells contribute to skin fibrosis. Sci Transl Med 10. [DOI] [PubMed] [Google Scholar]
- Thannickal VJ (2013). Mechanistic links between aging and lung fibrosis. Biogerontology 14, 609–615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todaro GJ, and Green H (1963). Quantitative studies of the growth of mouse embryo cells in culture and their development into established lines. J Cell Biol 17, 299–313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todaro GJ, Green H, and Goldberg BD (1964). Transformation of Properties of an Established Cell Line by Sv40 and Polyoma Virus. Proc Natl Acad Sci U S A 51, 66–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai CC, Wu SB, Kau HC, and Wei YH (2018). Essential role of connective tissue growth factor (CTGF) in transforming growth factor-beta1 (TGF-beta1)-induced myofibroblast transdifferentiation from Graves’ orbital fibroblasts. Sci Rep 8, 7276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tschumperlin DJ (2013). Fibroblasts and the ground they walk on. Physiology (Bethesda) 28, 380–390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner NA (2016). Inflammatory and fibrotic responses of cardiac fibroblasts to myocardial damage associated molecular patterns (DAMPs). J Mol Cell Cardiol 94, 189–200. [DOI] [PubMed] [Google Scholar]
- Uezumi A, Fukada S, Yamamoto N, Ikemoto-Uezumi M, Nakatani M, Morita M, Yamaguchi A, Yamada H, Nishino I, Hamada Y. et al. (2014). Identification and characterization of PDGFRalpha+ mesenchymal progenitors in human skeletal muscle. Cell Death Dis 5, e1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uezumi A, Fukada S, Yamamoto N, Takeda S, and Tsuchida K (2010). Mesenchymal progenitors distinct from satellite cells contribute to ectopic fat cell formation in skeletal muscle. Nat Cell Biol 12, 143–152. [DOI] [PubMed] [Google Scholar]
- Van Linthout S, Miteva K, and Tschope C (2014). Crosstalk between fibroblasts and inflammatory cells. Cardiovasc Res 102, 258–269. [DOI] [PubMed] [Google Scholar]
- Verrecchia F, Chu ML, and Mauviel A (2001). Identification of novel TGF-beta /Smad gene targets in dermal fibroblasts using a combined cDNA microarray/promoter transactivation approach. J Biol Chem 276, 17058–17062. [DOI] [PubMed] [Google Scholar]
- Virchow R (1858). Die Cellularpathologie in ihrer Begründung auf physiologische und pathologische Gewebelehre (A. Hirschwald). [PubMed]
- von Bubnoff A, and Cho KW (2001). Intracellular BMP signaling regulation in vertebrates: pathway or network? Dev Biol 239, 1–14. [DOI] [PubMed] [Google Scholar]
- Wang RN, Green J, Wang Z, Deng Y, Qiao M, Peabody M, Zhang Q, Ye J, Yan Z, Denduluri S. et al. (2014). Bone Morphogenetic Protein (BMP) signaling in development and human diseases. Genes Dis 1, 87–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei K, Korsunsky I, Marshall JL, Gao A, Watts GFM, Major T, Croft AP, Watts J, Blazar PE, Lange JK, et al. (2020). Notch signalling drives synovial fibroblast identity and arthritis pathology. Nature 582, 259–264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei P, Xie Y, Abel PW, Huang Y, Ma Q, Li L, Hao J, Wolff DW, Wei T, and Tu Y (2019). Transforming growth factor (TGF)-beta1-induced miR-133a inhibits myofibroblast differentiation and pulmonary fibrosis. Cell Death Dis 10, 670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werner S, and Alzheimer C (2006). Roles of activin in tissue repair, fibrosis, and inflammatory disease. Cytokine Growth Factor Rev 17, 157–171. [DOI] [PubMed] [Google Scholar]
- Wietecha MS, Pensalfini M, Cangkrama M, Muller B, Jin J, Brinckmann J, Mazza E, and Werner S (2020). Activin-mediated alterations of the fibroblast transcriptome and matrisome control the biomechanical properties of skin wounds. Nat Commun 11, 2604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkinson HN, Clowes C, Banyard KL, Matteuci P, Mace KA, and Hardman MJ (2019). Elevated Local Senescence in Diabetic Wound Healing Is Linked to Pathological Repair via CXCR2. J Invest Dermatol 139, 1171–1181 e1176. [DOI] [PubMed] [Google Scholar]
- Wipff PJ, Rifkin DB, Meister JJ, and Hinz B (2007). Myofibroblast contraction activates latent TGF-beta1 from the extracellular matrix. J Cell Biol 179, 1311–1323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie T, Liang J, Liu N, Huan C, Zhang Y, Liu W, Kumar M, Xiao R, D’Armiento J, Metzger D. et al. (2016). Transcription factor TBX4 regulates myofibroblast accumulation and lung fibrosis. J Clin Invest 126, 3626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie T, Wang Y, Deng N, Huang G, Taghavifar F, Geng Y, Liu N, Kulur V, Yao C, Chen P. et al. (2018). Single-Cell Deconvolution of Fibroblast Heterogeneity in Mouse Pulmonary Fibrosis. Cell Rep 22, 3625–3640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie Y, Su N, Yang J, Tan Q, Huang S, Jin M, Ni Z, Zhang B, Zhang D, Luo F. et al. (2020). FGF/FGFR signaling in health and disease. Signal Transduct Target Ther 5, 181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi Y, Itami S, Watabe H, Yasumoto K, Abdel-Malek ZA, Kubo T, Rouzaud F, Tanemura A, Yoshikawa K, and Hearing VJ (2004). Mesenchymal-epithelial interactions in the skin: increased expression of dickkopf1 by palmoplantar fibroblasts inhibits melanocyte growth and differentiation. J Cell Biol 165, 275–285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamakawa S, and Hayashida K (2019). Advances in surgical applications of growth factors for wound healing. Burns Trauma 7, 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang S, Cui H, Xie N, Icyuz M, Banerjee S, Antony VB, Abraham E, Thannickal VJ, and Liu G (2013). miR-145 regulates myofibroblast differentiation and lung fibrosis. FASEB J 27, 2382–2391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasuda M, Miyachi Y, Ishikawa O, and Takahashi K (2006). Spatial expressions of fibronectin and integrins by human and rodent dermal fibroblasts. Br J Dermatol 155, 522–531. [DOI] [PubMed] [Google Scholar]
- Zhang B, Tsai PC, Gonzalez-Celeiro M, Chung O, Boumard B, Perdigoto CN, Ezhkova E, and Hsu YC (2016). Hair follicles’ transit-amplifying cells govern concurrent dermal adipocyte production through Sonic Hedgehog. Genes Dev 30, 2325–2338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang LJ, Chen SX, Guerrero-Juarez CF, Li F, Tong Y, Liang Y, Liggins M, Chen X, Chen H, Li M. et al. (2019). Age-Related Loss of Innate Immune Antimicrobial Function of Dermal Fat Is Mediated by Transforming Growth Factor Beta. Immunity 50, 121–136 e125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang LJ, Guerrero-Juarez CF, Hata T, Bapat SP, Ramos R, Plikus MV, and Gallo RL (2015a). Innate immunity. Dermal adipocytes protect against invasive Staphylococcus aureus skin infection. Science 347, 67–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang W, Lavine KJ, Epelman S, Evans SA, Weinheimer CJ, Barger PM, and Mann DL (2015b). Necrotic myocardial cells release damage-associated molecular patterns that provoke fibroblast activation in vitro and trigger myocardial inflammation and fibrosis in vivo. J Am Heart Assoc 4, e001993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Tomann P, Andl T, Gallant NM, Huelsken J, Jerchow B, Birchmeier W, Paus R, Piccolo S, Mikkola ML, et al. (2009). Reciprocal requirements for EDA/EDAR/NF-kappaB and Wnt/beta-catenin signaling pathways in hair follicle induction. Dev Cell 17, 49–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang YE (2017). Non-Smad Signaling Pathways of the TGF-beta Family. Cold Spring Harb Perspect Biol 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Z, Kruglikov I, Zhao S, Zi Z, Gliniak CM, Li N, Wang MY, Zhu Q, Kusminski CM, and Scherer PE (2021). Dermal adipocytes contribute to the metabolic regulation of dermal fibroblasts. Exp Dermatol 30, 102–111. [DOI] [PubMed] [Google Scholar]
- Zhao J, Shi W, Wang YL, Chen H, Bringas P Jr., Datto MB, Frederick JP, Wang XF, and Warburton D (2002). Smad3 deficiency attenuates bleomycin-induced pulmonary fibrosis in mice. Am J Physiol Lung Cell Mol Physiol 282, L585–593. [DOI] [PubMed] [Google Scholar]
- Ziegler E (1895). General pathology; or, The science of the causes, nature and course of the pathological disturbances which occur in the living subject (New York: W. Wood and Company; ). [Google Scholar]
- Zwick RK, Guerrero-Juarez CF, Horsley V, and Plikus MV (2018). Anatomical, Physiological, and Functional Diversity of Adipose Tissue. Cell Metab 27, 68–83. [DOI] [PMC free article] [PubMed] [Google Scholar]