
Figure 1:
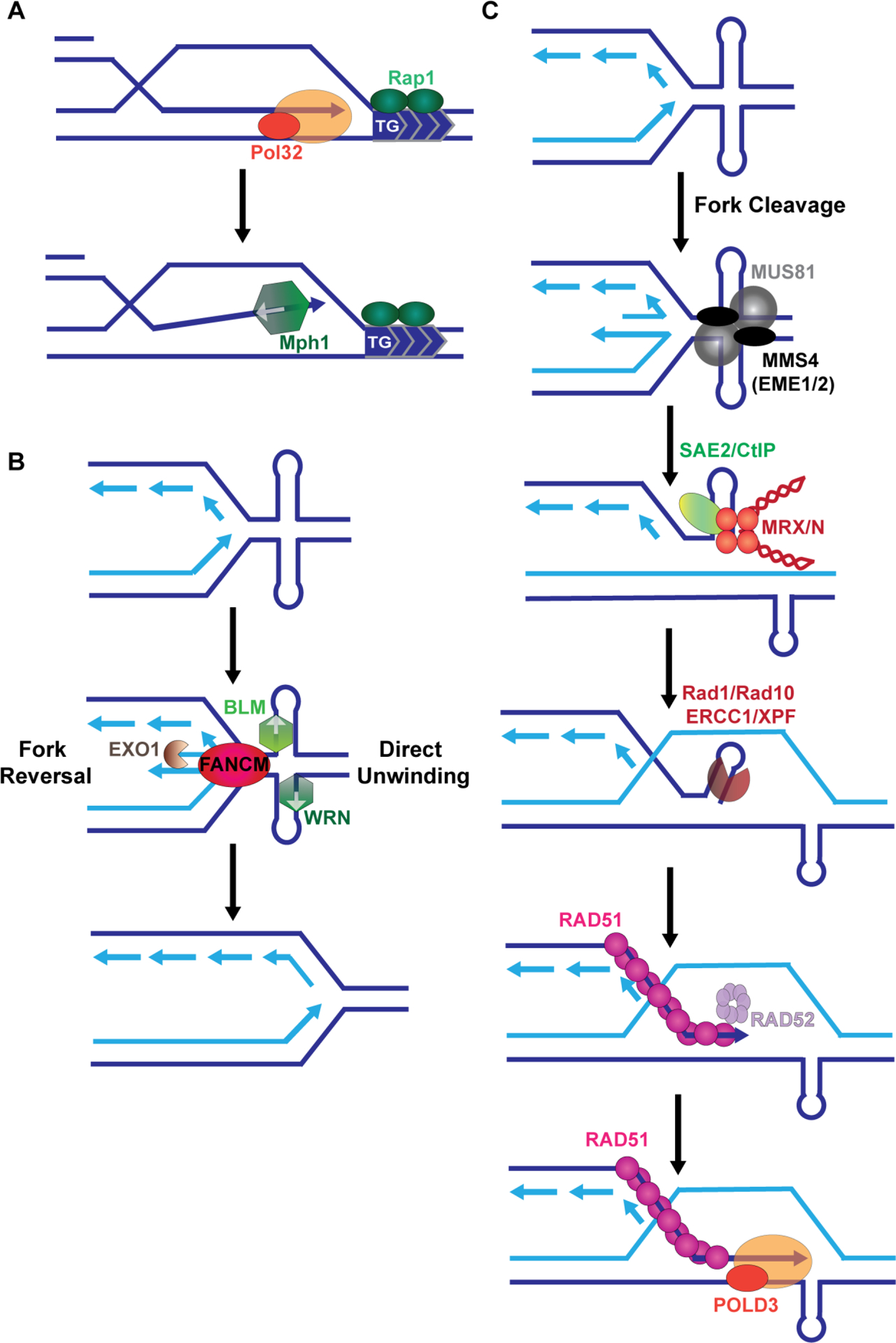
Models for replication fork restart and repair of collapsed forks at DNA structures. A. During BIR, if the replisome encounters a barrier, repair synthesis is more likely to abort. For internal telomeric sequences (ITS) native to that organism, de novo telomere addition on the recipient strand can lead to chromosome truncation. Abortive BIR due to the ITS is promoted by the helicase Mph1, which disassociates D-loops. B. DNA structures can stall replication, as illustrated for an AT repeat forming a cruciform structure. One mechanism for fork restart could be fork reversal which requires the translocase FANCM. Alternatively, helical unwinding of cruciform structures by the 3’−5’ helicases BLM or WRN can result in replication fork restart (note: other helicases are involved at G-rich hairpins or quadruplex structures, such as Srs2, Sgs1, RTEL and FANCJ [3]). C. In the event that fork restart fails, the replication fork may collapse. For some structures, breaks require cleavage by the MUS81-EME1 and/or the SLX1-SLX4 nucleases. The broken fork end with a hairpin is processed by MRX/N and Sae2/CtIP. Additional processing of the hairpin may occur by XPF/ERCC1 (Rad1/Rad10). Second end capture is mediated by RAD52 and repair synthesis is completed by Polδ with the accessory subunit POLD3.