
Figure 2:
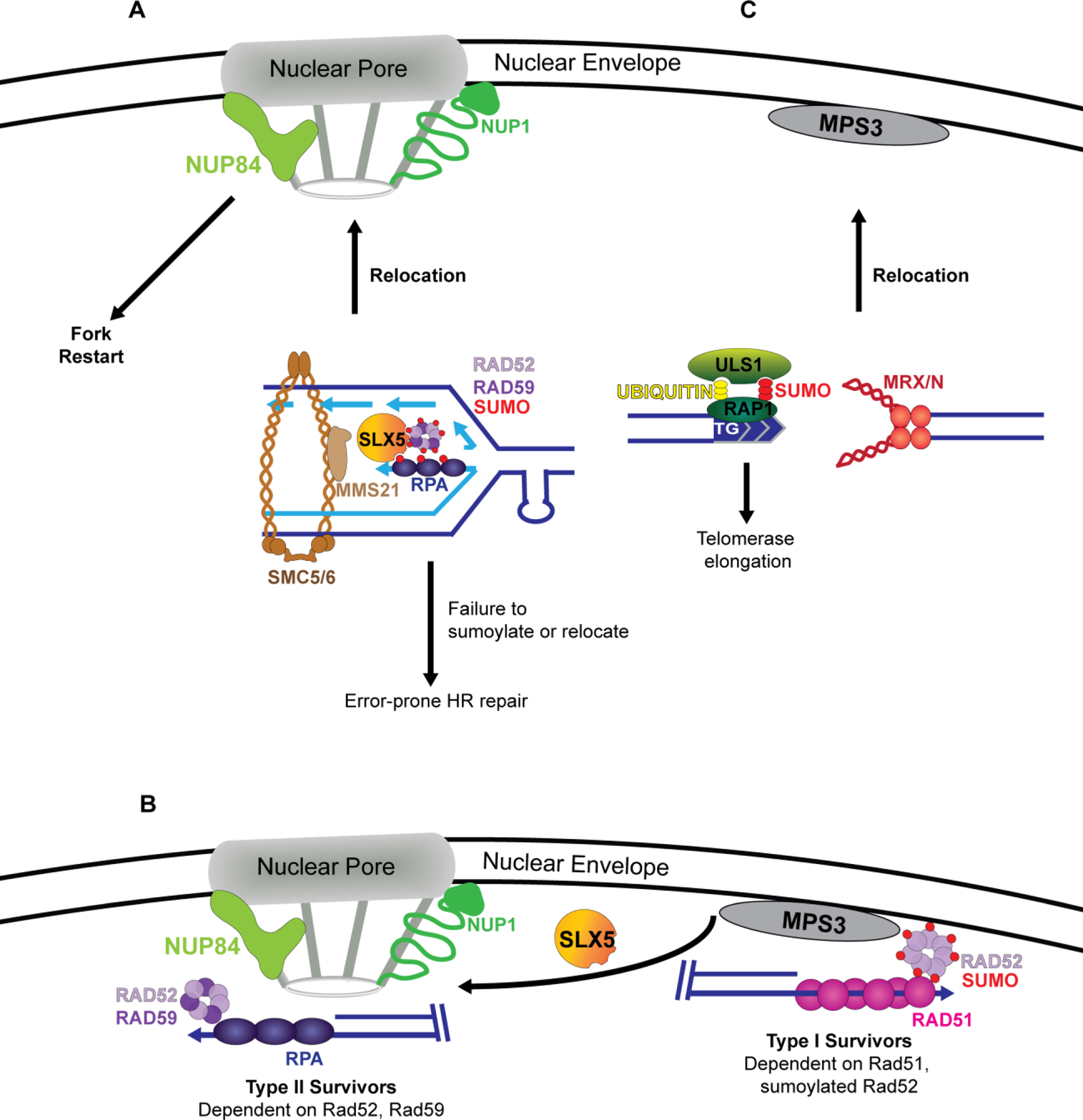
The role of nuclear location in controlling recombination and maintaining repetitive sequences. A. Replication forks stalled due to an expanded CAG repeat relocate to the nuclear pore complex (NPC) for repair and fork restart. Relocation is dependent on the StUbL Slx5/8 via Slx5 SIM domains and Mms21-mediated sumoylation of RPA, Rad52 and Rad59. Failure to sumoylate these proteins leads to impaired replication fork relocation, resulting in unconstrained recombination and repeat instability. B. Telomeres normally associate with the nuclear envelope. In the absence of telomerase, telomeres get progressively shorter until a crisis point, at which point they use recombination to extend the telomeric end. Eroded telomeres with sumoylated Rad52 bound result in type I survivors which depend on Rad51-mediated recombination. Further erosion results in relocation to the nuclear pore and the formation of type II survivors, which is dependent on Rad59 and de-sumoylation of Rad52. C. Targeted DSBs containing a TG repeat on one side of the break results in differential end processing. Broken ends that do not contain a telomeric seed are bound by MRX and relocate to the nuclear envelope. The end containing a telomeric seed is bound by the telomeric binding protein Rap1 which interacts with the StUbL Uls1, helping to signal telomerase recruitment to the TG sequence.