
Fig. 4.
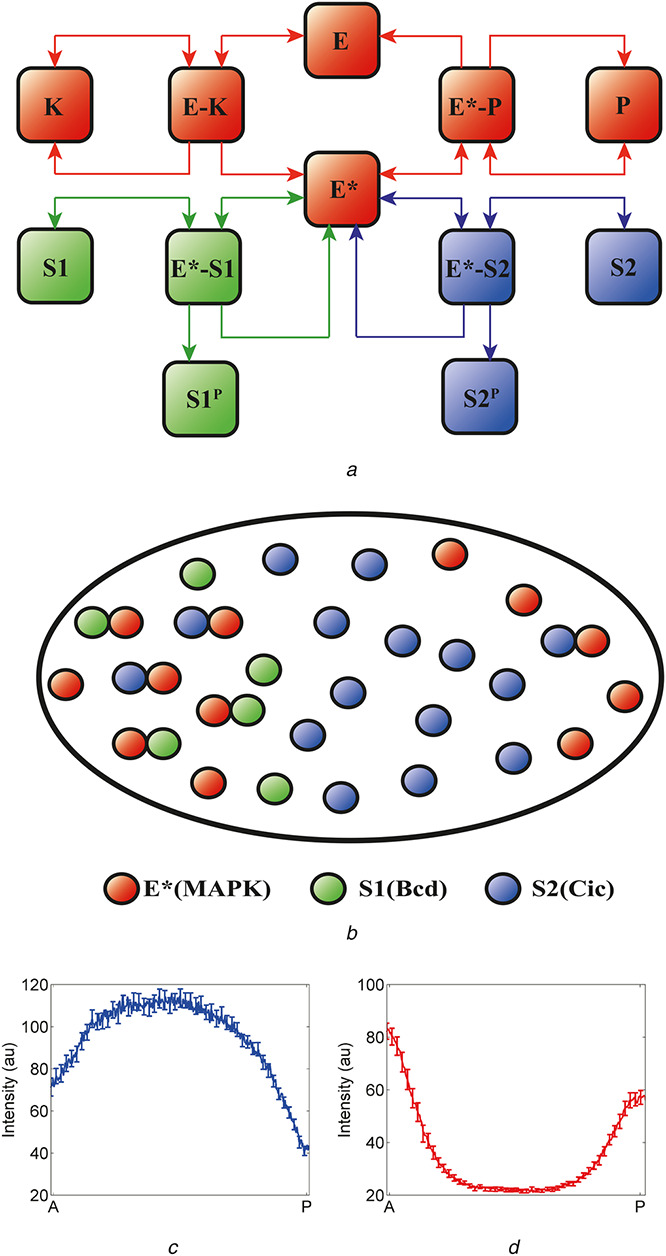
Substrate competition in the terminal patterning of the Drosophila embryo
(a) Substrate competition model: an enzyme (E) is activated when phosphorylated by a kinase (K). Activated enzyme (E*) then phosphorylates two substrates (S1 and S2) and is deactivated when dephosphorylated by a phosphatase (P). The two substrates compete both with each other and with the phosphatase for binding to the activated enzyme, (b) Schematic representation of the proposed substrate competition in the early Drosophila embryo. An enzyme is activated at the poles of the embryo (E*, red). At the anterior pole, there are both localised and uniform substrates, S1 (localised, green) and S2 (uniform, blue); at the posterior pole, there is only S2. In the context of the embryo, E* is phosphorylated MAPK, S1 models the combined effects of uniformly distributed substrates (such as Cic) and S2 models anteriorly localised substrates (such as Bcd), (c), (d) Non‐uniform spatial distribution of substrates leads to asymmetric pattern of phosphorylated MAPK and nuclear Cic. At the anterior pole, higher level of substrates provides effective protection of phosphorylated MAPK from the phosphatase, resulting in higher level of phosphorylated MAPK (d). At the same time, the level of Cic is also higher at the anterior pole due to the anteriorly localised substrates which act as competitive inhibitors of MAPK‐mediated downregulation of Cic (c)