
Fig. 6. Posterior Hox genes can support axial elongation.
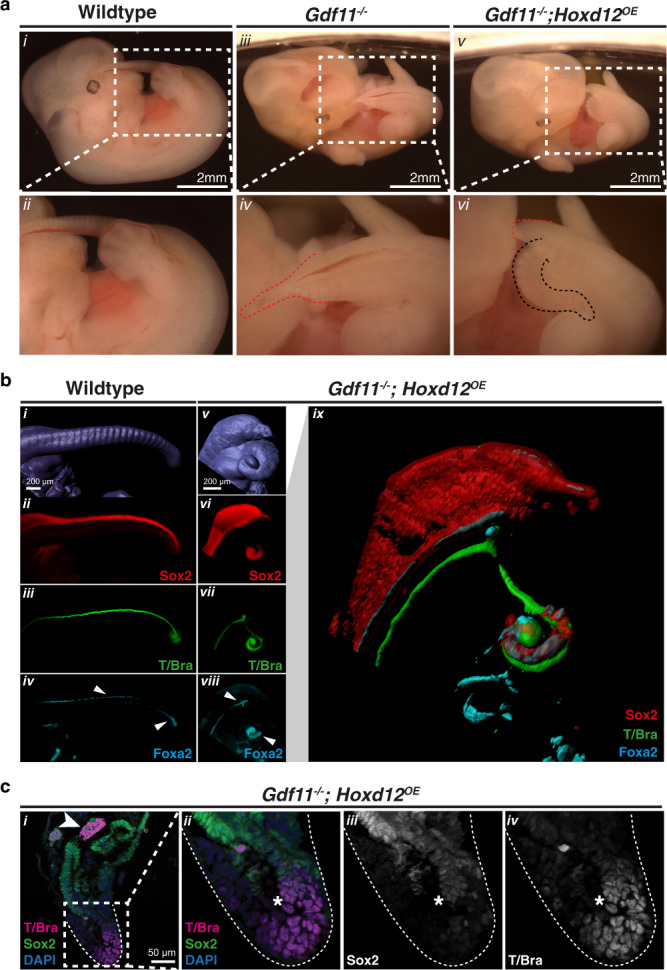
a Morphological analysis of tail phenotypes in E12.5 WT (i–ii), Gdf11−/− (iii–iv) and Gdf11−/−; Hoxd12OE (v–vi) mouse embryos. Brightfield images of whole embryos, with tail region enlarged for clarity. Red dotted line demarcates the caudal truncation phenotype observed in the majority of Gdf11−/− embryos, ending in a thinning blood-filled spike (iv) (n = 41/52). Black dotted line demarcates the ventral tail observed in the majority of Gdf11−/−;Hoxd12OE embryos (vi) (n = 22/32). b 3D reconstruction of caudal structures within E12.5 WT (i–iv) and Gdf11−/−; Hoxd12OE (v–ix) embryos immunostained for Sox2 (red), T/Brachyury (T/Bra; green) and Foxa2 (cyan). Enlarged and merged image of channels presented in (vi–viii) is shown in (ix). Tissue-restricted expression of Foxa2 within the ventral neural tube or tailgut is indicated using a white arrowhead. c Section immunofluorescence analysis reveals axial progenitor populations are present in the tailbud of Gdf11−/−;Hoxd12OE ventral tail at E12.5 (n = 2). Co-staining for T/Bra (magenta) and Sox2 (green) identified single-positive cells of either marker, along with dual positive cells indicative of neuromesodermal progenitors (asterisk). All sections are stained for DAPI (blue), arrowhead indicates notochord. n refers to the number of individual animals used for this analysis.