
Figure 2:
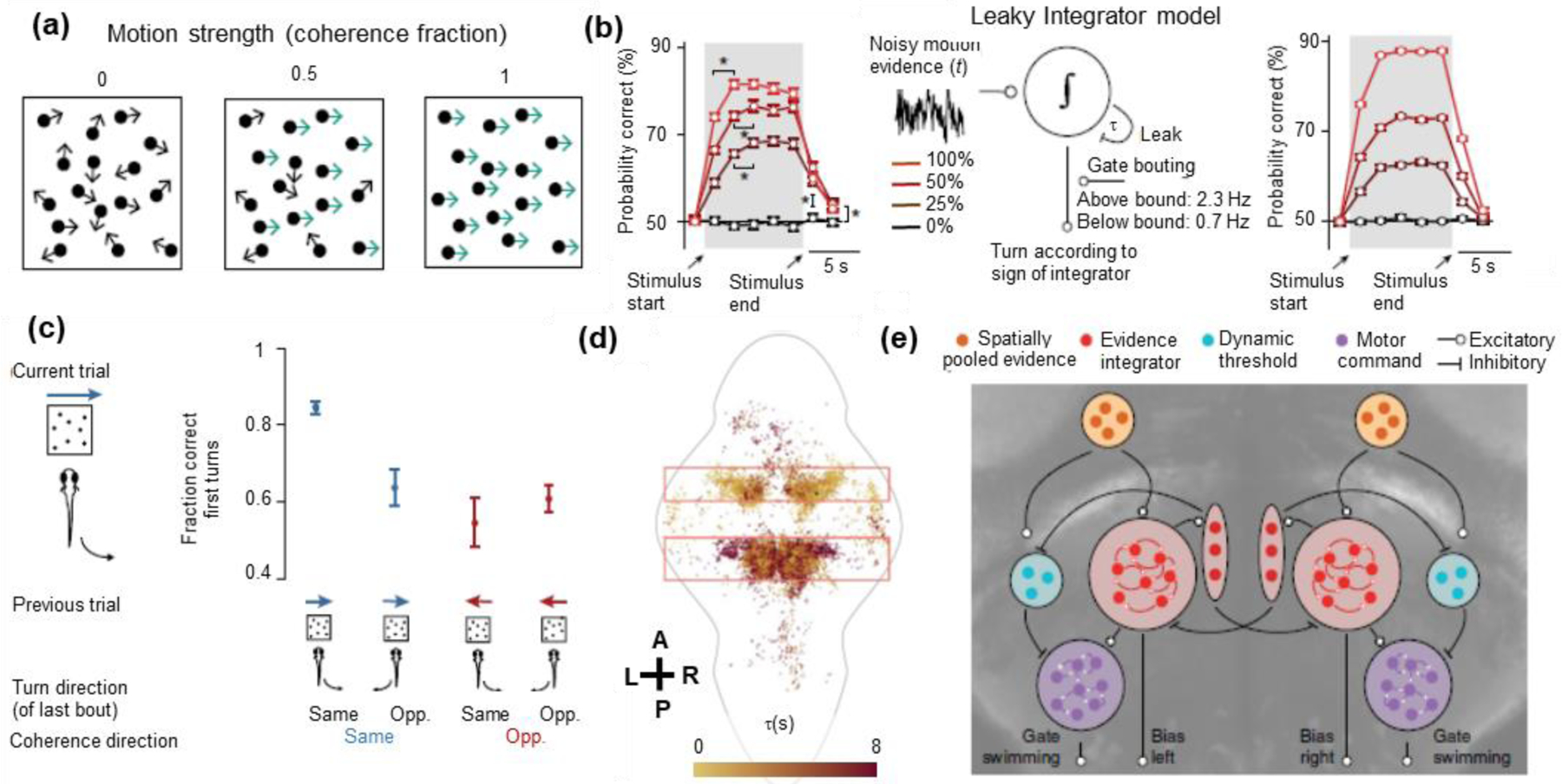
Mapping distributed sensorimotor processing in larval zebrafish using an evidence accumulation model. (a) Dragomir et al. [23] and Bahl et al. [24] used visual motion stimuli to drive turning behavior. Stimuli with three different strengths of rightward motion are shown (coherence fraction = 0, 0.5, 1). Panel from [23]. (b, left) The probability of correct turns (i.e. turns in the direction of motion) increased over time within the trial and also increased with higher coherence fractions. (b, middle) These results suggest an evidence accumulation model, whereby zebrafish leakily integrate noisy motion evidence to determine the direction of visual motion and set the rates of leftward and rightward turning. (b, right) The model successfully reproduces the observed behavioral patterns. Panel from [24]. (c) Motor actions do not reset the evidence accumulator, and its integration time scale is long enough to span several bouts and trials. The fraction of correct first turns accordingly depended on the direction of visual motion in the preceding trial and on the direction of the last turn. Panel from [23]. (d) Implementing the leaky integrator model would require the brain to first detect fast changes in sensory signals and then integrate them temporally. The authors therefore mapped brain activity by the integration time constant that best modeled each neuron’s stimulus responses. The map of model time constants showed a diversity of neuronal responses that were organized anatomically and could realize the computational steps of the behavioral models. Each colored dot is a segmented neuron, A-P denotes the anterior-posterior axis, and L-R denotes the left-right axis. Also shown are an outline of the zebrafish brain and boxes that highlight regions with many task-relevant neurons. Panel from [23]. (e) The authors ultimately superimposed their behavioral models onto cellular-resolution brain-wide maps, revealing neurons that detected motion signals in each direction, neurons that competitively pooled leftward and rightward motion signals, neurons that dynamically set the threshold separating low and high values of accumulated evidence, and neurons that represented stochastically generated leftward and rightward motor turning commands. Panel from [24].