
Fig. 3.
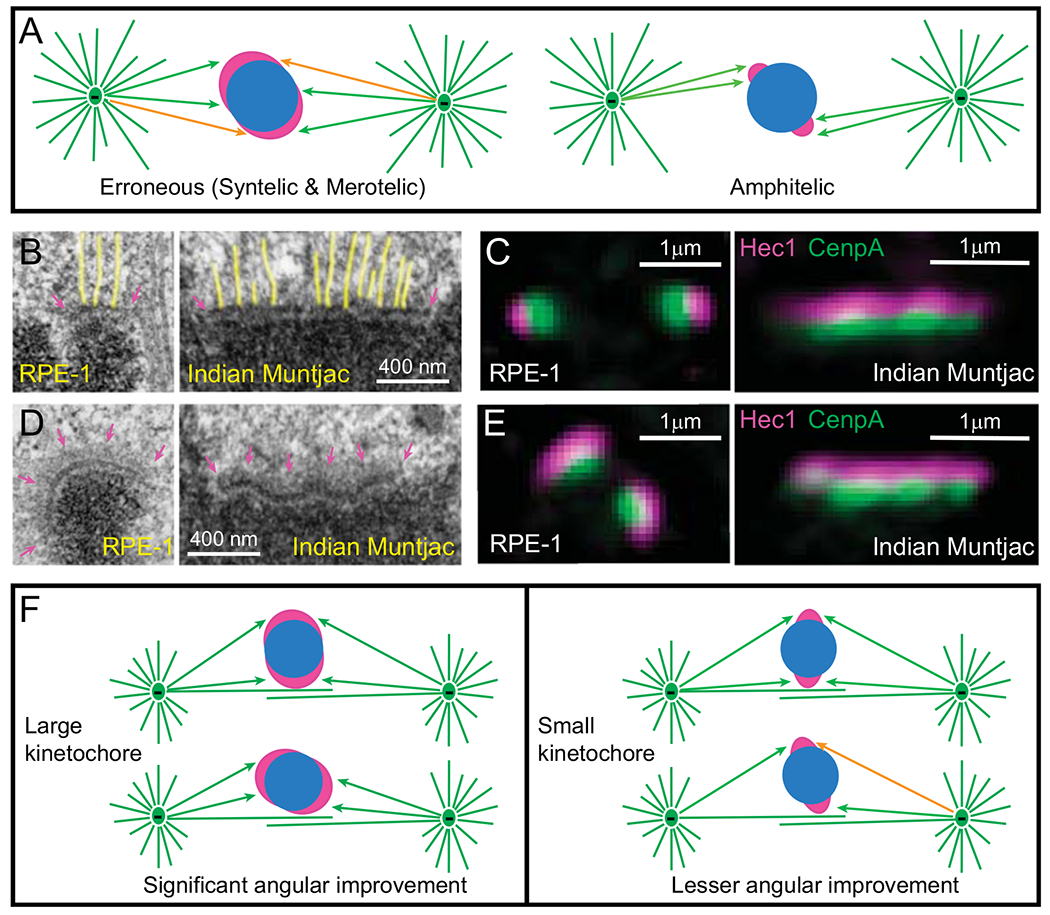
Effects of kinetochore architecture on the efficiency and fidelity of Search and Capture. (A) Reciprocity in the efficiency vs. fidelity of microtubule capture in the original S&C hypothesis. (Left cartoon) Larger kinetochores are likely to encounter microtubules faster; however, they are likely to be exposed to microtubules emanating from both spindle poles. This results in a mixture of proper (green arrows) and erroneous (orange arrows) interactions (syntelic and merotelic). (Right cartoon) Smaller kinetochores are protected from the exposure to ‘wrong’ spindle poles; however, microtubule search for small kinetochores is longer. (B–C) Architecture of attached kinetochores. (B) EM of metaphase kinetochores in human (RPE-1) and Indian muntjac cells. Arrows denote the trilaminar morphology of the plate comprising two electron-dense layers separated by a translucent middle layer. The length of the plate varies greatly. Notice end-on attached microtubules (highlighted yellow). (C) In fluorescence LM, mammalian kinetochores appear near diffraction-limited spots, while IM kinetochores form thin lines. Layered distribution of proteins within the kinetochore is apparent with DNA-binding proteins (CenpA-GFP, green) residing in the inner layer and microtubule-binding proteins (Hec1, magenta) in the outer layers. (D-E) Architecture of unattached kinetochores. (D) EM reveals fibrous corona (arrows in magenta) adjacent to the outer layer in the absence of microtubule attachments. In human cells (RPE-1), the plate elongates and encircles a large part of the centromere. In contrast, the length and shape of the plate does not change significantly in Indian muntjac. (E) Fluorescence LM is consistent with the elongation and shape changes in human (RPE-1) but not in Indian muntjac kinetochores. (F) Effects of kinetochore architecture on rotational alignment of the centromere in early prometaphase. Larger kinetochores support a higher degree of rotation on the surface of nascent spindle as they maintain constant contact with microtubules (left). In contrast, rotation angles of smaller kinetochores are sterically limited (right). As a result of extensive rotation, larger sister kinetochores have higher probability of forming proper attachments (green arrows) while less-aligned smaller kinetochores remain exposed to microtubules from ‘wrong’ spindle poles (orange arrow).