
Figure 4: Combining RNAi with short term auxin-mediated depletion.
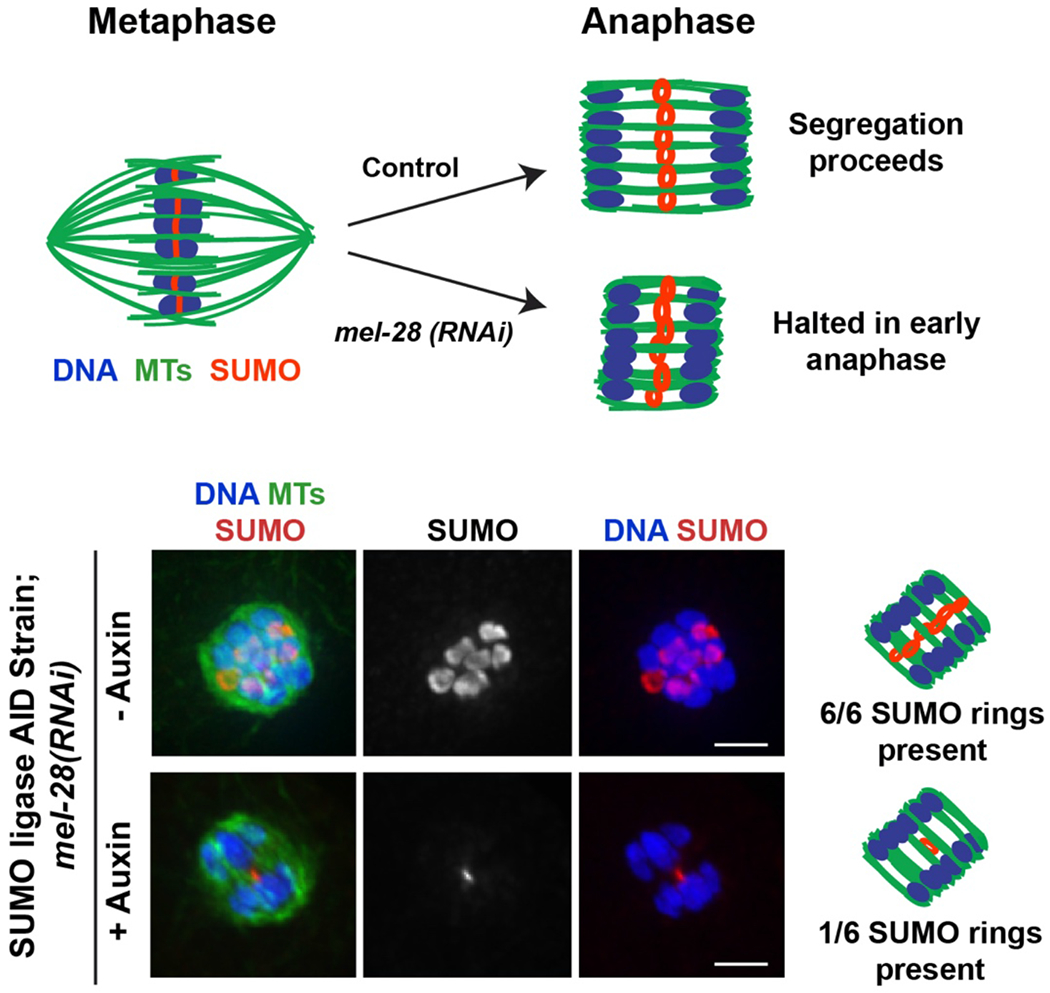
Studying the role of proteins during transient processes such as chromosome segregation is made easier by combining RNAi with the short term degron approach (Basic Protocol 2). As an example, we present depletion of a specific protein during anaphase in oocyte meiosis, modified from a recent study (Davis-Roca et al., 2018). In C. elegans oocytes, a ring complex with a modification known as SUMO (red) forms at the center of each of the six bivalents (blue) (Pelisch et al., 2017; Wignall & Villeneuve, 2009). In anaphase, these rings are left behind in the center of the spindle (green) (Davis-Roca et al., 2018; Davis-Roca et al., 2017; Dumont et al., 2010; Mullen & Wignall, 2017; Muscat et al., 2015). Knocking down the protein MEL-28 arrests spindles at early anaphase (Hattersley et al., 2016), making it possible to study the effects of depleting proteins at this particular stage (diagrams at top). In the example immunofluorescence images (bottom), short term auxin treatment was used to deplete the SUMO conjugating enzyme GEI-17 during this early anaphase arrest, induced by depleting MEL-28 using RNAi (protocol in (Davis-Roca et al., 2018)). MEL-28 depletion alone causes segregation to halt at a stage where all 6 SUMO rings are stabilized (top row of images). In contrast, 20 minutes of auxin treatment via soaking (to deplete GEI-17) results in fewer SUMOylated rings in the anaphase spindle. This suggests that the SUMO conjugating enzyme GEI-17 plays a role in stabilizing the ring complex during anaphase. Scale bar = 2μm.