
FIGURE 9.
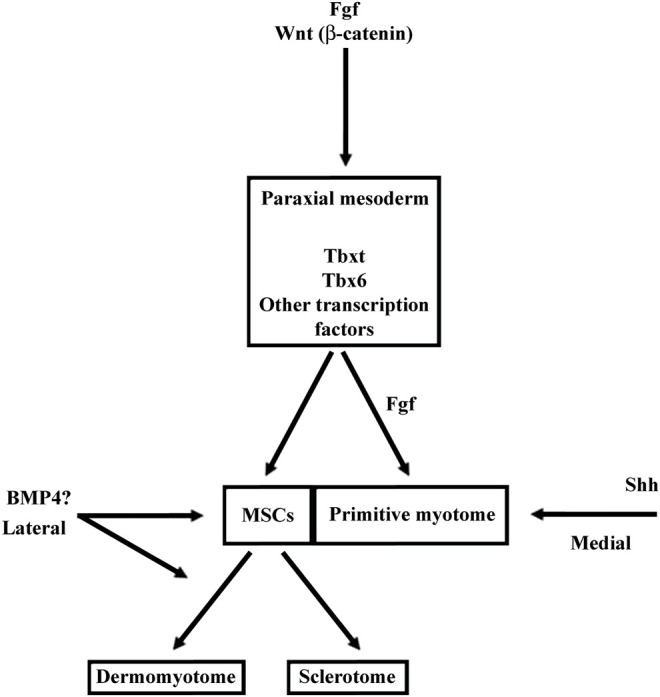
The signaling pathways involved in MSCs and primitive myotome formation in Xenopus. In anamniotes and particularly in Xenopus, the somite development is characterized by the early and massive myotome formation, and the delayed sclerotome development. The construction of primitive myotome is so early specified that it is interconnected to the dorso-mesoderm induction and the paraxial mesoderm specification. The signaling pathways like Fgf, Wnt and Nodal, involved in the dorso-mesoderm induction and in the paraxial mesoderm specification, also quickly trigger myogenic program leading to the primitive myotome formation (Jones et al., 1995; Wylie et al., 1996; Joseph and Melton, 1997; Fisher et al., 2002; Dorey and Amaya, 2010). In contrary, the amniote myotome formation takes place later after somitogenesis and the same signaling pathways involved earlier in the mesoderm induction and in the paraxial mesoderm specification did not induced myogenic program at the same time (Alev et al., 2013; Kiecker et al., 2016). In Xenopus, Fgf and Wnt play a key role in gene expression of the dorso-lateral marginal zone. This region can be considered as the presumptive paraxial mesoderm since it will give rise to somites later. Both Fgf and Wnt also contribute to the expression of Myf5 and Myod1 during the medial myogenic wave in Xenopus. Fgf has also been identified as the main inducer of the lateral myogenic wave which occurs later. In the beginning of neurulation, while the MSCs appear at the LSF, sonic hedgehog (Shh) secreted from notochord favors the myotome formation. BMP4 acts during neurulation to favor satellite cells lineage. Since the satellite cells are not already present at this stage, BMP4 would rather promote the MSCs and/or dermomyotome formation (Daughters et al., 2011).