
Figure 1.
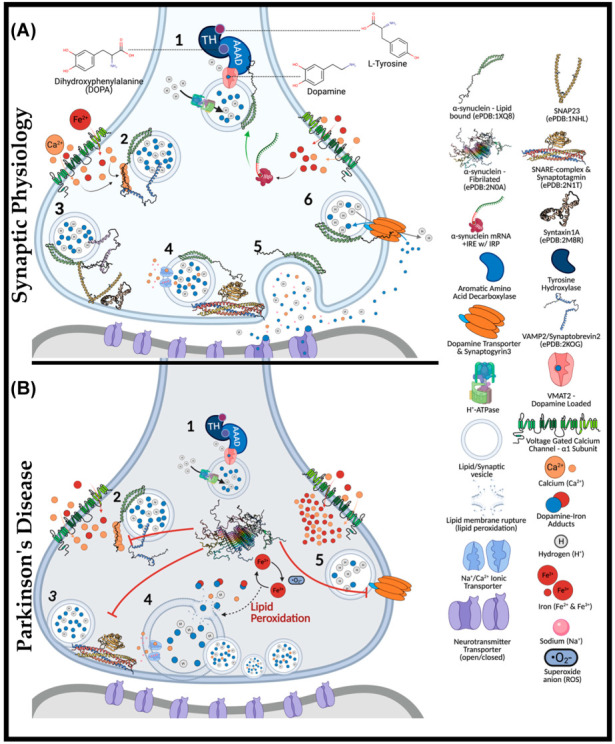
The Role of α-Synuclein at the Synaptic Junction in Physiology and in Parkinson’s Disease. In normal physiology (A): (1) complexed tyrosine hydroxylase (TH) and aromatic amino acid decarboxylase (AAAD) bind to the vesicular monoamine transporter (VMAT2), yielding vesicular dopamine. Vesicle membrane-bound H+-ATPases lower the intra-vesicular pH. Lipid-vesicle embedded α-synuclein has a role (2) in chaperoning soluble N-ethylmaleimide-sensitive factor attachment protein receptors (SNARE) after calcium activation. C-terminal binding between calcium-activated α-synuclein and vesicular-associated membrane protein 2 (VAMP2), exposes the SNARE motif for synaptosome-associated protein of 25 kDa (SNAP25) binding. SNAP25 recruits syntaxin-1A, and with the assistance of accessory proteins (synaptotagmin and Munc-18/13, an acronym for mammalian uncoordinated-18 or -13), the SNARE complex is formed (4), prompting the exocytosis of vesicular cargo (5). The α-synuclein is critical for synaptic plasticity by recycling synaptic vesicles post-exocytosis. Afterwards, (6) α-synuclein associates with the dopamine transporter to bring vesicles into proximity for synaptogyrin3 to modulate direct dopamine influx from the synaptic cleft. (B) (1) complexed tyrosine hydroxylase (TH) and aromatic amino acid decarboxylase (AAAD) bind to the vesicular monoamine transporter (VMAT2), yielding vesicular dopamine. Iron has also been suggested to permeate voltage-gated calcium channels (VGCCs), increasing the intracellular iron pool that may drive α-synuclein synthesis via a controversial and atypical 5′ iron-responsive element (IRE) in the untranslated region of α-synuclein mRNA. However, in the absence of lipid membrane vesicles, excessive iron-induced α-synuclein synthesis can yield disordered species that readily fibrillate (2) upon calcium exposure. The α-synuclein aggregates bind VAMP2 C-termini to block SNARE complex assembly (3). Consequently, vesicles cluster at the presynaptic membrane (4), while iron redox chemistry can lead to lipid peroxidation. Integral membrane damage can rupture vesicles, expelling reactive catecholamines, which lead to further damage through redox-active dopamine-iron adducts. Dopamine recycling is halted (5) as α-synuclein aggregates block vesicles docking at the dopamine transporter (DAT).