

Abstract
Background:
Bacterial vaginosis– a condition defined by a shift from Lactobacillus dominance to a polymicrobial, anaerobic bacterial community - increases the risk of acquiring sexually transmitted infections and other complications of the female reproductive tract. Antibiotic treatment frequently fails to return the microbiome to an optimal Lactobacillus-dominated state. No criteria currently exist to identify patients likely to experience treatment failure.
Objective(s):
We sought to identify pre-treatment community signatures associated with treatment failure through 16S rRNA gene analysis.
Study Design:
Twenty-eight women enrolled in an oral metronidazole treatment trial of BV were studied. Cervicovaginal lavage samples were collected prior to metronidazole treatment, and again at 7 and 30-days post-treatment. Cervicovaginal lavage DNA was amplified and sequenced using a paired-end, V4 region, 2×150 MiSeq run.
Results:
Of the 28 women, 25% failed to clear bacterial vaginosis, 35.7% demonstrated a transient clearance - shifting to community-type 2 (Lactobacillus iners dominant, CT2) at visit 2 only; 7.1% demonstrated delayed clearance - reaching CT2 at the final visit only; and 32.1% of patients experienced sustained bacterial vaginosis clearance. Examination of community composition and structure demonstrated that both richness and evenness were significantly lower for women who sustained clearance, while women who failed to clear bacterial vaginosis possessed the highest median levels of richness, evenness, and diversity pre-treatment. Soluble immune factors in the lower reproductive improved significantly following a shift from CT4 to a Lactobacillus-dominant microbiome, with samples categorized as CT2 possessing significantly higher levels of SLPI, GROa, and MIP3a and significantly lower levels of ICAM-1. While shifts to Lactobacillus-dominance improved markers of mucosal tissue health, these gains were only temporary amongst women who experienced recurrence.
Conclusions:
Assemblies of highly diverse microbiota are associated with the enhanced resilience of bacterial vaginosis to standard metronidazole treatment. These communities may be foundational to treatment resistance or simply an indication of a well-established community made possible by canonical biofilm forming taxa. Future studies must target the transcriptional activity of these communities under the pressure of antibiotic treatment to resolve the mechanisms of their resistance.
Keywords: Antibiotics, BV recurrence, molecular-BV, mucosal immunity, vaginal microbiome, biofilms
Introduction
Bacterial Vaginosis (BV) is a polymicrobial condition in the female reproductive tract (FRT) that is known to increase the risk of sexually transmitted infection (STI) acquisition and reproductive sequelae1–6. BV rates among women globally vary widely; recent estimates suggest a 30% prevalence in the US, while rates in sub-Saharan Africa may exceed 50%7,8. Despite this high prevalence, no single causative agent of BV is known.
The most universal feature of BV is the loss of Lactobacillus dominance in the vaginal ecosystem, which is accompanied by an increase in the proportion of polymicrobial anaerobic taxa8. The disruption of Lactobacillus dominance increases pH levels and nutrient availability, enabling the outgrowth of bacterial vaginosis-associated bacteria (BVAB) such as Gardnerella vaginalis14,15. Persistent colonization by Gardnerella vaginalis is associated with the development of biofilms that support other BVAB, including Atopobium, Prevotella, Sneathia, and Shuttleworthia16,17,18,19.
The largescale availability of marker gene sequencing data has enabled the stratification of vaginal communities into discrete categories defined by their composition – frequently referred to as cervicotype (CT)1 or community state types (CSTs)13. Dominance by Lactobacillus crispatus (CT1) is considered an optimal state and is linked to a marked decrease in STI susceptibility. CT2 is dominated by Lactobacillus iners and is frequently considered a transitional phenotype to CT3 and CT4 - collectively referred to as “molecular-BV”14,15. These communities, which are dominated by Gardnerella vaginalis or a highly diverse BVAB population (CT3 and CT4, respectively), strongly correlate with an increase of inflammatory factors, reproductive sequelae, and STI susceptibility1–6.
Despite these known risks, BV recurrence rates remain above 70% within 12 months of treatment22. Current CDC guidelines recommend metronidazole as a first line treatment23. As a pro-drug, metronidazole is reduced to its active state under anaerobic conditions – a mechanism that spares beneficial Lactobacillus. However, the complete removal of BVAB is likely complicated by the presence of biofilms, which shield BVAB by preventing metronidazole penetration.
Here, we build upon an examination of women with symptomatic BV who enrolled in a clinical study (CONRAD 115, ClinicalTrials.gov NCT01347632)6. The initial analysis identified several BV-associated changes; these included increased inflammatory mediators and HIV target cells in the FRT, and reduced antimicrobial activity of the CVL6. We hypothesized that 16S ribosomal RNA (rRNA) gene analysis would identify keystone taxa or community structures that were present at diagnosis and that contribute to BV recurrence or treatment failure; our primary objective was to leverage these data toward the development of a molecular predictor of treatment failure.
Materials and Methods
Human and nonhuman experimentation: enrollment and sampling
The longitudinal, open-label study was approved by the Chesapeake Institutional Review Board (IRB) (Pro #00006122) with a waiver of oversight from the Eastern Virginia Medical School and was registered in ClinicalTrials.gov (#NCT01347632). Thirty-three women seeking treatment for symptomatic vaginal discharge with or without atypical vaginal odor were enrolled upon Nugent scoring of 4 or higher; Amsel criteria were not utilized in this study. All participants were non-pregnant, free of reproductive tract infections, and denied use of antibiotics within the last 14 days. Upon enrollment, vaginal swabs, cervico-vaginal tissue biopsies and cervicovaginal lavage (CVL) were collected. Following collection of biological samples, all women were prescribed fourteen 500mg metronidazole tablets, with one tablet to be taken twice daily; patient adherence was self-reported. Biological samples were collected again between 7 and 10 days following the conclusion of treatment (visit 2), and again 28–32 days post-treatment (visit 3). Biological samples included vaginal pH testing, and CVL was collected through administration of 10cc of normal saline. Three full thickness cervico-vaginal biopsies were obtained under topical anesthesia at the conclusion of CVL collection.
Soluble factor analysis
CVL SLPI was measured by ELISA (R&D Systems, Minneapolis, MN) using a victor2 reader (Perkin Elmer Life Sciences, Boston, MA). Interleukin (IL)-1b, IL-6, IL-8, IL- 10, ICAM-1, and tumor necrosis factor alpha were measured in undiluted CVLs by a multiplex electro-chemiluminescence assay (Meso Scale Discovery, Gaithersburg, MD). All measurements were performed in duplicate.
DNA extraction
Bacteria from cervicovaginal lavage samples were pelleted by centrifugation and resuspended in 200μl of lysis buffer (30mM Tris-HCl, 10mM EDTA, 200mM sucrose, pH 8.2). Samples were heated at 65°C for 10 minutes prior to the addition of 100mg/ml lysozyme solution to a final concentration of 10mg/ml. Samples were then incubated for 1 hour at 37°C. 5% SDS was then added to a final concentration of 1% w/v and incubated at 56°C for 10 minutes. DNA extractions were then performed using Qiagen’s DNeasy Blood and Tissue Kit.
16S rRNA sequencing
Extracted DNA was amplified following the Earth Microbiome Protocol for 16S Illumina sequencing utilizing 515F-806R primers originally described by Caporaso et al.34 to target the V3–V4 region of the 16S SSU rRNA. This broadly-applicable – and widely utilized - primer set was chosen to maximize the comparability of this data set with existing and future marker gene sequencing of the vaginal microbiome. Amplicon concentrations were normalized, pooled, and cleaned prior to KAPA quantification. The pooled library was sequenced using a 2×150 bp Illumina MiSeq run.
Metataxonomic analyses
16S rRNA amplicon sequencing reads were demultiplexed in Illumina Basespace. A demultiplexed read object was generated in qiime2-2018.2. Amplicon Sequence Variants (ASVs) were generated using the dada2 denoise-single with 0 trim-left and trunc at 145. Phylogeny was determined using mafft-fastree option and ASVs were assigned taxonomy using the 99% Greengenes 515–806 classifier. Qiime2 objects were exported for downstream analysis in R. All fastq files were deposited in SRA under ascension PRJNA691964.
Results
Lactobacillus Recovery Varies Following Metronidazole Treatment
Twenty-eight of 33 women were confirmed to have molecular BV by 16S rRNA sequencing, while five patients found to have Lactobacillus-dominant microbiomes pre-treatment were removed from further analyses. At diagnosis, mean Lactobacillus abundance was 4.4% (Fig. 1a,e). Known BVAB, such as Shuttleworthia, Sneathia, Prevotella, and Gardnerella, were the most likely to be the dominant genus in a given sample at baseline (Fig. S1a,b). Several other highly prevalent bacterial genera frequently associated with BV diagnosis were observed at mean abundances of less than 5%, including Megasphaera, Atopobium, Clostridium, Parvimonas, Dialister, Peptonophilus, and Gemella (Fig. 1d,e).
Figure 1 –
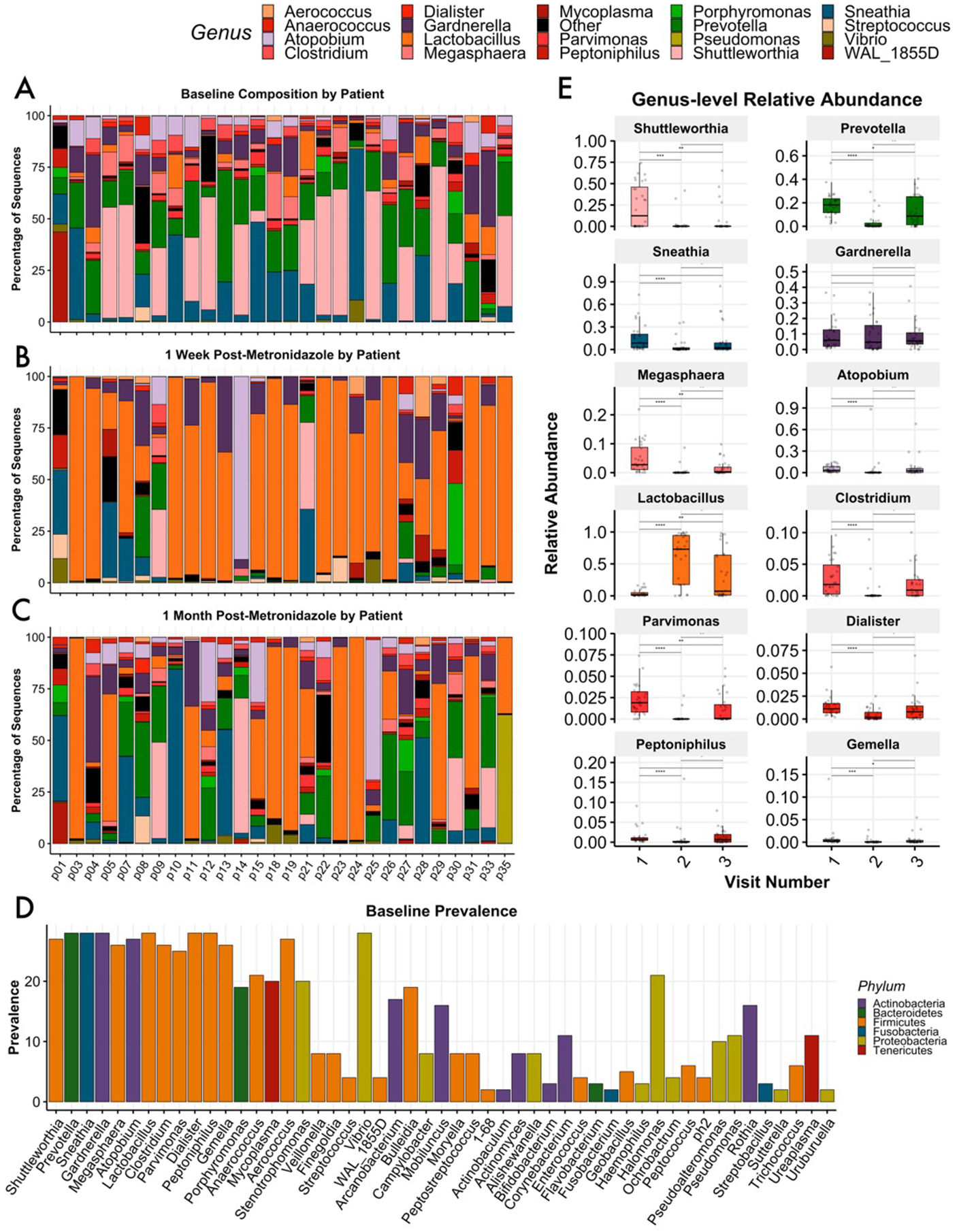
Baseline and post-treatment vaginal microbiota profiles of women diagnosed with Nugent-BV. Relative abundance taxonomic plots of cervicovaginal lavage assessed by 16S rRNA gene profiling; women were sampled at baseline upon BV diagnosis (A), 7 – 10 days after conclusion of 10 day course of metronidazole (B), and again at 1 month post-treatment (C). A histogram demonstrates the majority of genera considered BV-associated were universally prevalent at baseline, including Prevotella, Sneathia, Gardnerella,Lactobacillus, Dialister, and Peptonophilus; Shuttleworthia and Atopobium were present in 27 of 28 analyzed samples. Several rare taxa were also highly prevalent (D). Box-and-whisker plots define median and interquartile ranges; all BV-associated bacteria deteceted in this study were significantly reduced at visit 2 following metronidazole treatment, though mean levels roseagain at visit 3. Lactobacillus was significantly increased at visits 2 and 3 (E).
At visit 2 one-week post-treatment, Lactobacillus mean relative abundance reached 70.1%, while several genera that were dominant at baseline decreased significantly (Fig. 1b,e). At visit 3, approximately one-month post-treatment, mean Lactobacillus abundance was down to 24.7% across all samples, while Sneathia, Prevotella, and Shuttleworthia increased on average (Fig. 1c,e). Mean values were found to obscure divergent treatment outcomes; these are discussed in detail below.
The alpha diversity measurements of richness and evenness assess the number of observed taxa per sample and the degree of parity amongst taxa, respectively, while inverse Simpson suggests the complexity of a sample by estimating the likelihood that two randomly chosen reads from a sample would identify the same taxa. In the context of BV treatment, positive therapeutic responses include reducing the observed diversity and increasing dominance by Lactobacillus. While all measurements of diversity were significantly decreased at the level of the cohort for visit 2, a number of individuals actually experienced increases in richness and diversity (Fig. 2a). Further analysis revealed that a subset of women who did develop Lactobacillus iners dominance by visit 2 failed to retain these communities at visit 3, indicating BV recurrence. The variable response to treatment can be measured by Beta diversity, which estimates the degree off difference between two communities; these differences can be visualized using dimensionality reduction techniques such as Principal Coordinates Analysis (PCoA), where an increasing distance between any two samples represents decreasing similarity. In the present study, while patients clustered together at baseline, community composition and structure varied widely following treatment (Fig. 2b). It should be noted that patients were not found to cluster by age, BMI, cycle day, contraception method, or self-reported race(Fig.S6a–e).
Figure 2 –
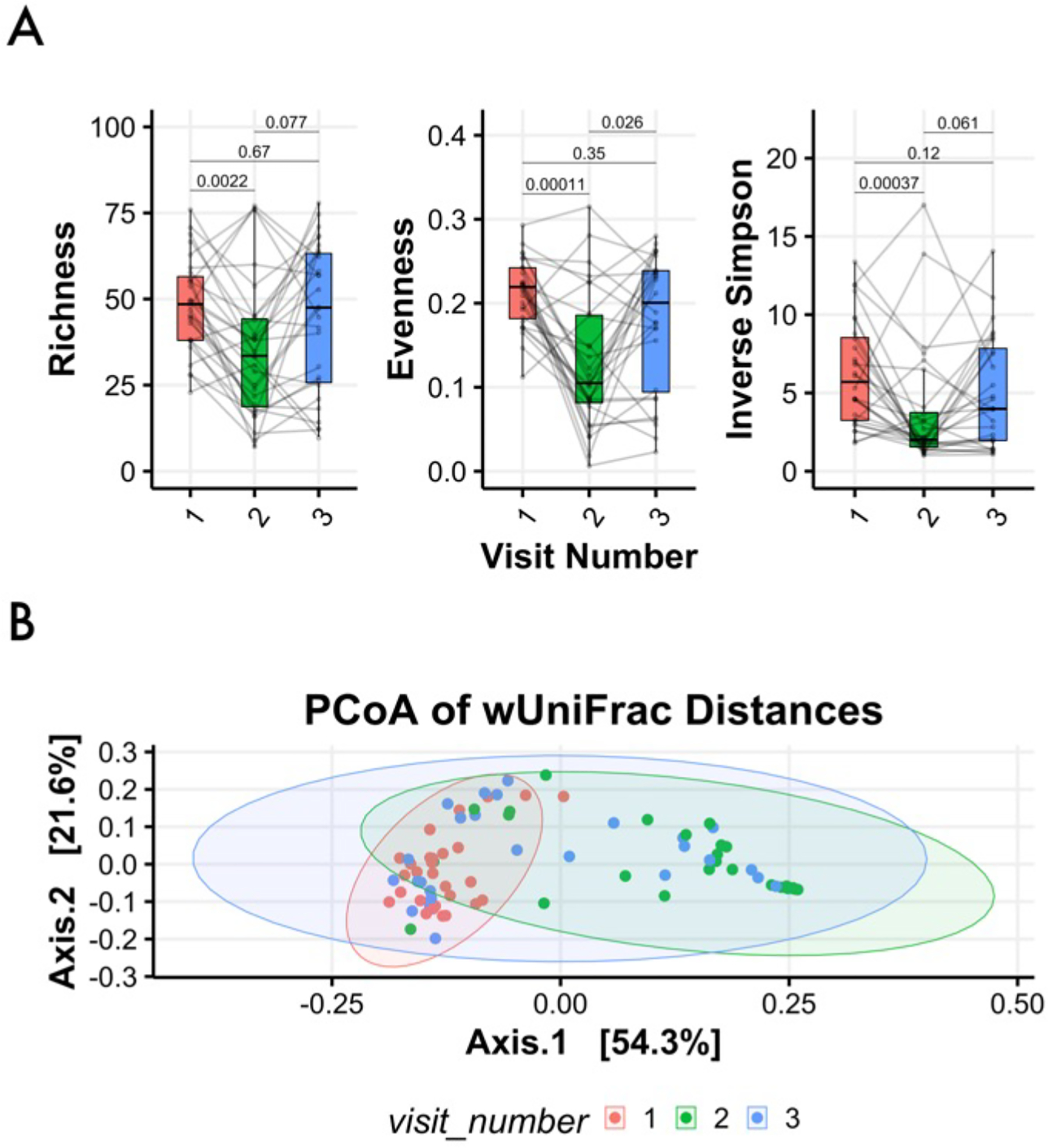
Alpha and Beta Diversity assessment. Box-and-whisker plots define median and interquartile ranges for measures of alpha diversity such as richness, evenness, and the inverse Simpson index, which were significantly decreased one week post-treatment; only evenness remained significantly reduced at visit 3 (A). A principal coordinate analysis of weighted UniFrac distances with samples (dots) colored here by visit: visit one (red), two (green), and three (blue); shaded ellipses represent 95% confidence intervals for cervicotype distribution. The plot illustrates samples clustered at visit 1 prior to treatment, while samples were considerably more varied at visits 2 and 3, reflecting divergent responses to metronidazole therapy (B).
Soluble Immune Factors in FRT Respond to Microbiota Shifts
Given the diverse patterns of change in microbial structure following treatment for BV, we cervicotyped patients at each visit. We elected to use the classification strategy described in Anahtar et al., which utilizes 4 CTs assigned according to dominant organism, with CT1 dominated by Lactobacillus crispatus, CT2 by Lactobacillus iners, CT3 by Gardnerella vaginalis, and CT4 by any of a number of diverse anaerobes associated with BV. We first validated the distinct composition of CTs through wUNIFRAC assessment (Fig. 3a), finding significant differences between each CT pairwise (2v3: adj.p=0.003,2v4: adj.p=0.003,3v4:adj.p=0.027); CT1 was not observed in the current study. We next assessed vaginal pH levels across these groupings and found that women supporting CT2 had a significantly lower pH at the time of sampling than women in either CT3 and CT4 (Fig. 3b; p=0.001; p=3.9e-12). In addition to differences in vaginal pH, we identified significantly higher levels of SLPI, GROa, and MIP3a in CVL from women in CT2 as compared to CT4(Fig. 3c); these women also had significantly lower levels of ICAM-1 (p=2e-4) and a trend toward lower IL-1B (p=0.067). CD4+ cells from the lamina propria as assessed by microscopy were significantly higher in women categorized as CT4(p=0.034), in agreement with recent reports4,6.
Figure 3 –
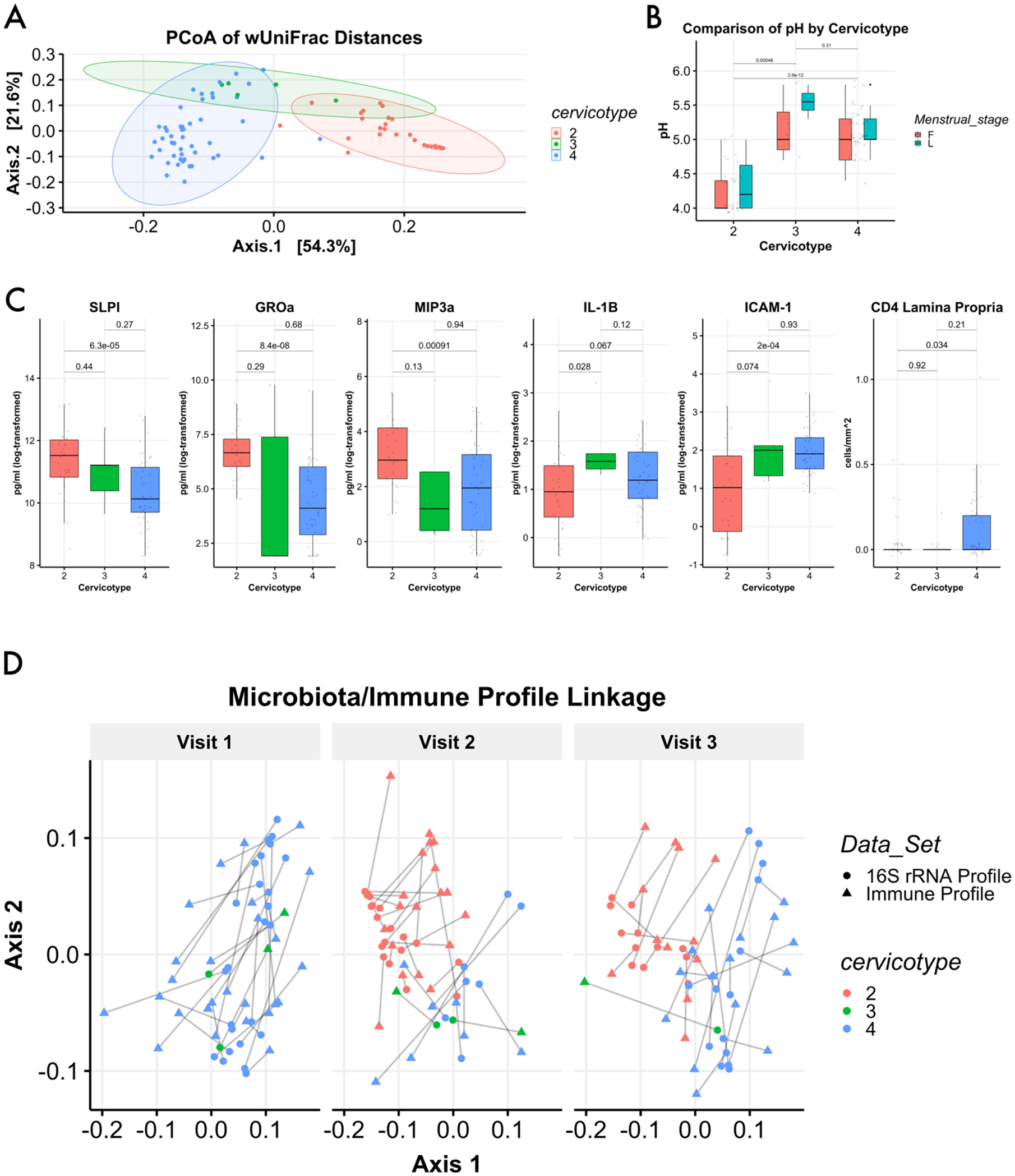
Immune profiles segregate by cervicotype. Principal coordinate analysis based on weighted UniFrac distances demonstrated clustering of samples by cervictoype. Samples (dots) are colored here by cerivcotype: CT2 in red; CT3 in green, and CT4 in blue. CT1 was not observed in this study. Shaded ellipses represent 95% confidence intervals for cervicotype distribution (A). Assessment of sample pH based on assigned cervicotype shows significant elevations in pH for women with cervicotype 3 or 4 (B). Several soluble factors analyzed from cervicovaginal lavage showed signficant differences between CTs 2, 3, and 4; additionally, CD4+ cells in the lamina propria were significantly elevated for women in CT4 (C). Vaginal immune profiles were generated through measurement of select soluble factors present in CVL. Samples were then compared based on the aggregated measurements through Bray-Curtis dissimilarity and visualized through principal coordinate analysis, with the plot split into visits 1, 2, and 3. 16S rRNA gene data was visualized in the same plot through procrustes rotation. By visit 2, women who transititioned to Lactobacillus dominance possessed a statistically distinct immune profile (p.adjusted = 0.003 by permanova), and this distinction remained significant at visit 3 one-month post-treatment (p.adjusted = 0.003 by permanova). The emergence of a distinct immune profile for women with Lactobacillus dominance is paralleled by the emergence of distinct bacterial community compositions associated with a shift to CT2 (p.adjusted = 0.006 at visit 2 and visit 3 by permanova) (D).
We hypothesized that immune profiles generated by combining all measured soluble factors (Table S1) would demonstrate an alignment between CT and immune response for each sample collected. Dimensionality reduction of the log-transformed values demonstrated that immune profiles clustered by CT (Fig. S3A; adj.p=0.003). Importantly, the clusters generated through immune profile analysis strongly resemble clusters generated using metataxonomic data (Fig. S3B). When combined using Procrustes rotation - a technique to compare ordinations by minimizing the residual sum of squares between plots - these data demonstrate that a shift in CT was consistently associated with a shift in immune profile (Fig. 3d).
Community Structure at Treatment Initiation Impacts Response to Metronidazole
Assignment of CT for each patient by visit identified patients who experienced failed, transient, delayed, or sustained BV clearance as determined by 16S rRNA gene analysis. Twenty-five percent of patients failed to clear BV (did not improve to CT2 at any visit); 35.7% demonstrated a transient BV clearance - shifting to CT2 at visit 2 only; 7.1% demonstrated delayed clearance - reaching CT2 at the final visit only; and 32.1% of patients experienced sustained BV clearance - maintaining a shift to CT2 at both visit 2 and 3. Both the transient and sustained BV clearance groups experienced significant community reorganization following metronidazole treatment; specifically, Lactobacillus relative abundance swelled at visit 2 (Fig. 4a), with reciprocal decreases to Shuttleworthia, Prevotella, Sneathia, and Atopobium (Fig. 4b). For women that failed to clear BV, relative abundance of BV-associated taxa such as Shuttleworthia and Prevotella were moderately reduced at visit 2, while Atopobium levels were increased. In cases of transient BV clearance, BVAB levels at visit 3 returned to - or exceeded – levels detected at BV diagnosis.
Figure 4 –
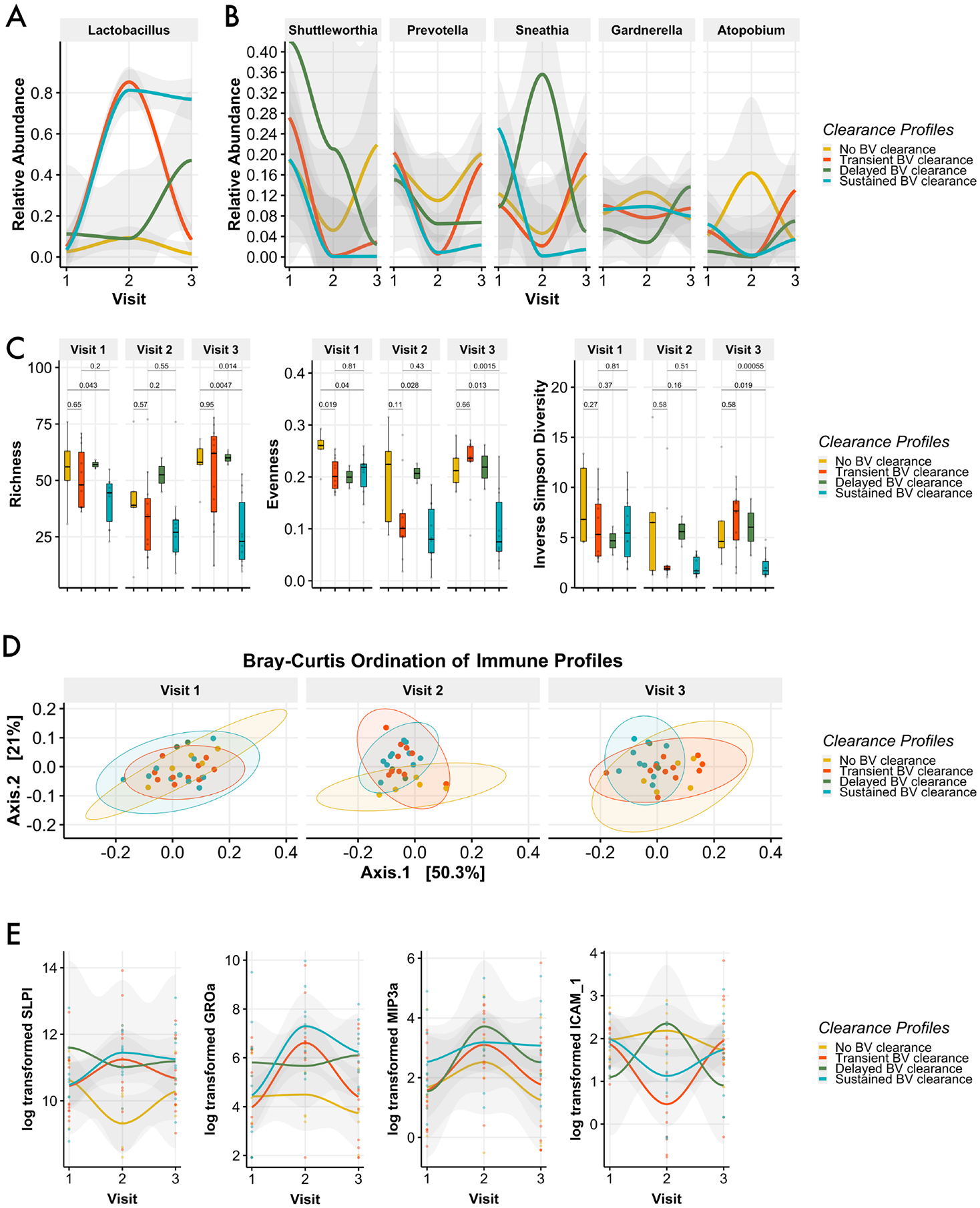
Post-treatment shifts to Lactobacillus-dominance vary in timing and duration. Only a portion of women experiened significant increases in Lactobacillus levels following treatment; of these women who were cleared of SeqBV at visit 2, only a portion remained clear at visit 3 (A). Relative abundance levels of several BV-associated bacteria seen here were inversely related to gains in Lactobacillus; levels of these bacteria were at or above baseline level for many women at the final visit (B). Women who sustained SeqBV clearance following treatment had significantly lower levels of observed taxa and measured evenness prior to treatment initiation (C). SeqBV clearance patterns were not predicated by immune profile at baseline (D). Smoothed plot of select soluble factors over time that were shown to differ significantly by CT (E).
To evaluate our hypothesis that elements of the community structure at the time of treatment initiation impact outcome, we first assessed distinctions in alpha diversity metrics between clearance profiles. Women who failed to clear BV demonstrated the highest median levels of richness, evenness, and diversity at visit 1 (Fig. 4c). Importantly, both richness and evenness were significantly lower for women that sustained BV clearance versus those women who failed to clear BV (Fig 4c). All groups were indistinguishable at baseline by beta diversity assessment(Fig. S4), and this finding suggests that the accumulation of rare or low-abundance taxa may play a larger role than dominant taxa in determining treatment outcome. Accordingly, analysis of differential abundance to identify keystone taxa that were significantly enriched or notably absent failed to identify meaningful pre-treatment differences between any of the 4 clearance groups. Finally, we asked if the occurrence or duration of CT shifts following metronidazole treatment could be due to an immunological predisposition and found that immune profiles for the 4 response groups were indistinguishable at baseline (Fig. 4d). Importantly, analyses of post-treatment visits demonstrated distinct patterns of change that stratified according to clearance profile (Fig. 4e).
Comment
Principal findings
Here, a significant increase in the prevalence of a diverse collection of low-abundance taxa was associated with metronidazole treatment failure. Equally important, women who experienced total treatment failure or BV recurrence experienced a subsequent increase in prevalence of these taxa above their baseline levels. While markers of mucosal health improved with a shift to Lactobacillus dominance, these changes were transient in women who experienced recurrence.
Results in the Context of What is Known
In this study, we stratified metronidazole treatment outcomes according to the occurrence and duration of vaginal microbiota shifts to Lactobacillus-dominated CT2. Importantly, not a single woman in this study shifted to CT1, the most optimal vaginal microbial structure. This approach also allowed us to ask if community characteristics prior to treatment initiation contributed to treatment outcome. We found that the microbiota of women with a durable shift to Lactobacillus-dominance were significantly less complex at diagnosis than those women who failed to shift to CT2. These findings suggest that metronidazole treatment is more likely to be effective when community richness is lower. As niche space is subdivided within an ecosystem, functional resistance to environmental stressors is often increased26,27, and thus, the development of a high complexity vaginal microbiota may increase compositional stability. Indeed, research into the stability of vaginal microbiota across health and disease has demonstrated that women who shift to a higher, more complex CT are unlikely to return to a lower complexity state28.
Within the context of antibiotic pressure, highly diverse communities may benefit polymicrobial anaerobic assemblies by providing the collective capacity to interfere with environmental metronidazole activity; this notion is strongly supported by a growing body of research that has demonstrated a role for both gut and vaginal microbes to metabolize, alter, and sequester drugs29,30,31. Alternatively, highly diverse communities may simply reflect well-established scaffolds known to play a role in relapsing BV; specifically, biofilms produced by genera such as Gardnerella vaginalis provide barriers to antibiotics and can serve to protect other taxa as well; Atopobium specifically have been shown to comprise as much as 40% of BV-associated biofilms17. Given the significant role that community richness may play in treatment failure, it is important to note that women in this study who experienced transient shifts to CT2 following treatment concluded the study with elevations in mean diversity, not reductions. These data suggest that ineffective treatment may predispose women to future treatment failure and facilitate development of more intractable BV. Indeed, we found that in cases of treatment failure or relapse, the biofilm-associated genera Atopobium and Sneathia generally increased after antibiotic exposure(Fig. 4b), a finding in agreement with research that has shown Atopobium to be a strong contributor to BV relapse34.
In addition to these findings, our work adds to the growing body of evidence demonstrating microbiota-induced modification of antimicrobial responses and immunity in the vaginal mucosa. Our finding that immune profiles clustered by CT at the time of sampling suggests that patients undergoing treatment for BV experience microbiome-mediated immune shifts as community composition changes. This strongly supports the growing body of work that indicates a direct role of the microbiome in modifying mucosal immune health in the FRT1. These immune shifts were likely advantageous for women who shifted to Lactobacillus-dominated CT2, as they experienced significant increases in levels of SLPI -– which is both antimicrobial and known to protect mucosal barrier tissues from damage caused by neutrophil elastase6. Importantly, increased levels of SLPI are associated with reduced risk of HIV acquisition34. Women who shifted to CT2 also demonstrated a significant reduction in ICAM-1 – a biomarker for FRT inflammation6 – which was further supported by a trend toward reduced IL-1B, a strongly pro-inflammatory cytokine6.
Clinical Implications
Taken together, our data demonstrate that treatment failure or BV recurrence following metronidazole therapy may actually strengthen factors associated with intractable BV. Treating all patients diagnosed with BV as a single, homogenous condition may place women at risk. Sequence-based assessments that would identify these established, resistant communities are currently not accessible in the clinical setting. This indicates an intense need to develop bedside tools capable of staging BV prior to treatment. It should be noted that while Nugent score was positively correlated with microbiota richness by all metrics (Fig. S5a), there was some discordance between patients deemed BV+ by Nugent score and those categorized as BV- by sequencing analysis(Fig. S5b).
The conspicuous finding that none of the women observed here shifted to CT1, which is dominated by Lactobacillus crispatus, suggests that ongoing efforts to develop biotherapeutics that supply active cultures of L. crispatus may be an essential aspect of future treatment. Several ongoing safety and efficacy trials should be monitored closely for clinically viable therapeutics.
Research Implications
While presence/absence approaches such as 16S rRNA gene analysis and shotgun metagenomics have provided novel insights to the complex makeup of the vaginal microbiome, understanding of the functional activities of the frequently unculturable taxa it contains should be sought in the form of transcription-based assays. Because DNA-based approaches utilize genomic material that may persist following treatment, early timepoints may identify taxa as present despite their death due to treatment. Additionally, these assays fail to provide mechanistic responses to therapy; the identification of transcriptional changes in response to metronidazole could provide insights into community vulnerabilities that translate into new therapeutic approaches.
Strengths and Limitations
A major strength of our assessment is that it provides previously unavailable insights into factors associated with metronidazole treatment failure. Several previous studies have investigated the most dominant BVAB through PCR-based methods and found that their levels do not determine therapeutic outcomes. Here, our unbiased approach allowed us to identify that increased diversity of “minor” BVAB may play a larger role in treatment outcomes than previously expected. Additionally, our work expands upon previous cross-sectional studies by showing that improvements to mucosal immune health fluctuate in real time with improvements in microbiota composition – and that these changes are transient when BV recurs.
Caveats include an absence of measurements for antibiotic levels, though all patients self-reported taking metronidazole as prescribed. Future studies would be improved through the collection of BV recurrence data; this could enable a comparative assessment of microbial communities between women experiencing their first episode of BV treatment and those women who have been treated for BV previously. While a larger sample size in a similar microbiome marker study would lend additional strength to our findings, it is notable that the data collected from the 28 enrolled participants were sufficient to discriminate between cervicotypes and response profiles - and ultimately detect significant differences in (a) microbiota complexity prior to treatment and (b)mucosal immune profiles as community profiles shifted. Additionally, while the sample size of 28 is modest, the ability to track the sequenced-microbiome prior to and following therapeutic treatment with antibiotics is a major strength of the work – and one that is currently lacking in the literature.
Additionally, shotgun sequencing approaches would enhance the ability to detect antimicrobial resistance (AMR) genes – a study that is lacking in the field at this time. Finally, keystone bacteria such as Lactobacillus iners are known to alter transcriptional profiles substantially in the context of increased diversity; a metatranscriptional approach could resolve how bacterial responses stratify according to treatment outcome.
Conclusions
Current approaches to managing BV do not adequately assess the risk of treatment failure, which is demonstrated by the unacceptably high levels of relapse experienced by women receiving the standard of care. The presence or absence of individual taxa seems unlikely to be a direct contributor to treatment outcome, meaning clinicians continue to lack a prognostic biomarker. Our data suggest complex community dynamics govern the collective response to treatment. Therefore, while one arm of research must aim to provide clinicians with bedside staging strategies that anticipate difficult-to-treat cases, another must examine the mechanistic underpinnings of BV persistence toward the goal of providing them with targeted therapeutics.
Supplementary Material
Supplemental Figure 1 – Dominant taxa at Baseline. Histogram breaking down the number of samples dominated by specific genera (A). Relative abundance of the dominant genus in each sample (B).
Supplemental Figure 2 – Longitudinal Lactobacillus levels. Levels of Lactobacillus tracked by patient over the course of the study.
Supplemental Figure 3 – immune Profile Relationship to 16 rRNA gene profile. Immune profiles generated from select soluble factors were assessed by Bray-Curtis dissimilarity and findings were visualized through PCoA; samples clustered by metataxonomic CT and facted by visit(A). Bray-Curtis analysis of CT classifications, facted by visit (B).
Supplemental Figure 4 – Baseline and follow up Beta diversity clustered by clearance profile. Bray-Curtis analysis of women from each clearance profile, faceted by visit. (A).
Supplemental Figure 5 – Alpha diversity metrics of richness, Shannon diversity, and inverse simpson diversity show positive correlation with Nugent score (A). Discordance between Nugent BV scores and and seqBV are visualized by the number of patients in each cervicotypes. Colors indicate the dominant taxa in each sample (B).
Supplemental Figure 6 – PCoA visualization of wUNIFRAC analysis of women by age, BMI, menstrual cycle day and stage, birth control strategy and self-reported race. (A).
Supplemental Table 1 – Patient characteristics. Adapted from Thurman et al.
Supplemental Table 2 – Soluble factors. Soluble factors from CVL were measured as described in Methods.
Condensation:
Prevalence of low-abundance taxa are significantly elevated in women experiencing bacterial vaginosis treatment failure, suggesting diversification of established communities may entrench dominant,suboptimal bacteria.
AJOG at a glance:
-
Why was this study conducted?
Current approaches to managing bacterial vaginosis (BV) do not adequately assess the risk of treatment failure, which is demonstrated by the unacceptably high levels of relapse. This study sought to define aspects of microbiota composition at the time of BV treatment initiation that predispose women to treatment failure.
- What are the key findings?
- While levels of the most common BV associated bacteria do not predict risk of treatment failure, the accumulation of diverse, low-abundance taxa may predispose women toward treatment resistant BV.
- Shifts to Lactobacillus-dominance improved markers of mucosal tissue health, but these gains were only temporary amongst women who experienced recurrence.
What does this study add to what is already known?
Our study provides evidence that a highly diversified vaginal ecosystem leads to BV treatment failure, which concerningly, lead to increased diversity above baseline levels. The failure of metronidazole therapy could entrench existing polymicrobial communities.
Acknowledgements:
We would like to thank the entire study team of CONRAD, and all participants.
Sources of Funding:
These studies were supported through funding to NRK via R01DK112254 from NIDDK/NIH, MG via R01 AI145296 and R01 AI127463, and MA via AI138718 01. Additional funding mechanisms include the CONRAD 115 BV study was funded by an inter-agency agreement between the Centers for Disease Control and Prevention (CDC) and the United States Agency for International Development (USAID) through a cooperative agreement with CONRAD/EVMS (GPO-A-00-08-00005-00). The views of the authors do not necessarily represent those of the funding agencies; the funding agencies played no role in submission decision.
Footnotes
Clinical trial: NCT01347632, Registered April 14, 2011, clinicaltrials.gov, https://www.clinicaltrials.gov/ct2/show/NCT01347632, study start date: May 2012.
Will individual participant data be available (including data dictionaries)? No; Summary data are available from the publication of a previous manuscript: doi:10.1089/aid.2015.0006.
What data in particular will be shared? Patient demographics include: average age, BMI and years of education; self-reported race/ethnicity; contraception/sexual history
What other documents will be available (e.g., study protocol, statistical analysis plan, etc.)? The study protocol, inclusion/exclusion criteria, analyses plan, and primary and secondary objectives are available at https://www.clinicaltrials.gov/ct2/show/NCT01347632. A related manuscript (doi: 10.1089/aid.2015.0006) contains summary patient information during the study period including: day of menstrual cycle, vaginal pH, Antiviral and antibacterial capacity of the cervicovaginal secretions, Semiquantitative vaginal flora assessments, select cytokines and chemokines (pg/ml): IL-1B, Il-6, Il-8, IL-10, ICAM-1,TNF-a,SLPI,GRO-a,MIP-3a,RANTES, Elafin
When will data be available (start and end dates)? All data are currently – and indefinitely – available.
How will data be shared (including with whom, for what types of analyses, and by what mechanism)? These data are publicly available within the clinical trial registration, the NCBI-held manuscript and using the SRA bioproject id PRJNA691964.
Ethics approval and consent to participate: The longitudinal, open-label study was approved by the Chesapeake Institutional Review Board (IRB) (Pro #00006122) with a waiver of oversight from the Eastern Virginia Medical School (EVMS) and was registered in ClinicalTrials.gov (#NCT01347632).
Competing interests: The authors declare that they have no competing interests.
Disclosures: The authors report no conflict of interest.
Availability of data and material:
The datasets generated and/or analyzed during the current study are available in the Sequence Read Archive repository under ascension PRJNA691964.
References
- 1.Anahtar MN, Byrne EH, Doherty KE, et al. Cervicovaginal bacteria are a major modulator of host inflammatory responses in the female genital tract. Immunity. 2015. May 19;42(5):965–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Brotman RM, Klebanoff MA, Nansel TR, Yu KF, Andrews WW, Zhang J, Schwebke JR. Bacterial vaginosis assessed by gram stain and diminished colonization resistance to incident gonococcal, chlamydial, and trichomonal genital infection. Journal of Infectious Diseases. 2010. Dec 15;202(12):1907–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cherpes TL, Wiesenfeld HC, Melan MA, et al. The associations between pelvic inflammatory disease, Trichomonas vaginalis infection, and positive herpes simplex virus type 2 serology. Sexually transmitted diseases. 2006. Dec 1;33(12):747–52. [DOI] [PubMed] [Google Scholar]
- 4.Gosmann C, Anahtar MN, Handley SA, et al. Lactobacillus-deficient cervicovaginal bacterial communities are associated with increased HIV acquisition in young South African women. Immunity. 2017. Jan 17;46(1):29–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lennard K, Dabee S, Barnabas SL, et al. Microbial composition predicts genital tract inflammation and persistent bacterial vaginosis in South African adolescent females. Infection and immunity. 2018. Jan 1;86(1):e00410–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Thurman AR, Kimble T, Herold B,et al. Bacterial vaginosis and subclinical markers of genital tract inflammation and mucosal immunity. AIDS research and human retroviruses. 2015. Nov 1;31(11):1139–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kenyon C, Colebunders R, Crucitti T. The global epidemiology of bacterial vaginosis: a systematic review. American journal of obstetrics and gynecology. 2013. Dec 1;209(6):505–23. [DOI] [PubMed] [Google Scholar]
- 8.McKinnon LR, Achilles SL, Bradshaw CS, et al. The evolving facets of bacterial vaginosis: implications for HIV transmission. AIDS research and human retroviruses. 2019. Mar 1;35(3):219–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mitchell C, Marrazzo J. Bacterial vaginosis and the cervicovaginal immune response. American Journal of Reproductive Immunology. 2014. Jun;71(6):555–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Amsel R, Totten PA, Spiegel CA, Chen KC, Eschenbach D, Holmes KK. Nonspecific vaginitis: diagnostic criteria and microbial and epidemiologic associations. The American journal of medicine. 1983. Jan 1;74(1):14–22. [DOI] [PubMed] [Google Scholar]
- 11.Nugent RP, Krohn MA, Hillier SL. Reliability of diagnosing bacterial vaginosis is improved by a standardized method of gram stain interpretation. Journal of clinical microbiology. 1991. Feb;29(2):297–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Leitich H, Kiss H. Asymptomatic bacterial vaginosis and intermediate flora as risk factors for adverse pregnancy outcome. Best practice & research Clinical obstetrics & gynaecology. 2007. Jun 1;21(3):375–90. [DOI] [PubMed] [Google Scholar]
- 13.Ravel J, Gajer P, Abdo Z, Schneider GM, et al. Vaginal microbiome of reproductive-age women. Proceedings of the National Academy of Sciences. 2011. Mar 15;108(Supplement 1):4680–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gajer P, Brotman RM, Bai G, et al. Temporal dynamics of the human vaginal microbiota. Science translational medicine. 2012. May 2;4(132):132ra52–. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Srinivasan S, Liu C, Mitchell CM, et al. Temporal variability of human vaginal bacteria and relationship with bacterial vaginosis. PloS one. 2010. Apr 15;5(4):e10197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Marrs CN, Knobel SM, Zhu WQ, Sweet SD, Chaudhry AR, Alcendor DJ. Evidence for Gardnerella vaginalis uptake and internalization by squamous vaginal epithelial cells: implications for the pathogenesis of bacterial vaginosis. Microbes and infection. 2012. Jun 1;14(6):500–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Swidsinski A, Mendling W, Loening-Baucke V, et al. An adherent Gardnerella vaginalis biofilm persists on the vaginal epithelium after standard therapy with oral metronidazole. American journal of obstetrics and gynecology. 2008. Jan 1;198(1):97–e1. [DOI] [PubMed] [Google Scholar]
- 18.Swidsinski A, Verstraelen H, Loening-Baucke V, Swidsinski S, Mendling W, Halwani Z. Presence of a polymicrobial endometrial biofilm in patients with bacterial vaginosis. PloS one. 2013. Jan 8;8(1):e53997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Verstraelen H, Swidsinski A. The biofilm in bacterial vaginosis: implications for epidemiology, diagnosis and treatment. Current opinion in infectious diseases. 2013. Feb 1;26(1):86–9. [DOI] [PubMed] [Google Scholar]
- 20.Wessels JM, Lajoie J, Vitali D, et al. Association of high-risk sexual behaviour with diversity of the vaginal microbiota and abundance of Lactobacillus. PLoS One. 2017. Nov 2;12(11):e0187612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vitali D, Wessels JM, Kaushic C. Role of sex hormones and the vaginal microbiome in susceptibility and mucosal immunity to HIV-1 in the female genital tract. AIDS research and therapy. 2017. Dec;14(1):1–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bostwick DG, Woody J, Hunt C, Budd W. Antimicrobial resistance genes and modelling of treatment failure in bacterial vaginosis: clinical study of 289 symptomatic women. Journal of medical microbiology. 2016. May 1;65(5):377–86. [DOI] [PubMed] [Google Scholar]
- 23.Centers for Disease Control and Prevention. (2020, November 28). 2015 Sexually Transmitted Diseases Treatment Guidelines. Retrieved from https://www.cdc.gov/std/tg2015/bv.html
- 24.National Center for Biotechnology Information. (2021, January 10). Stat Pearls: Metronidazole. Retrieved from https://www.ncbi.nlm.nih.gov/books/NBK539728/
- 25.Mayer BT, Srinivasan S, Fiedler TL, Marrazzo JM, Fredricks DN, Schiffer JT. Rapid and profound shifts in the vaginal microbiota following antibiotic treatment for bacterial vaginosis. The Journal of infectious diseases. 2015. Sep 1;212(5):793–802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Baert JM, De Laender F, Sabbe K, Janssen CR. Biodiversity increases functional and compositional resistance, but decreases resilience in phytoplankton communities. Ecology. 2016. Dec;97(12):3433–40. [DOI] [PubMed] [Google Scholar]
- 27.Pennekamp F, Pontarp M, Tabi A, et al. Biodiversity increases and decreases ecosystem stability. Nature. 2018. Nov;563(7729):109–12. [DOI] [PubMed] [Google Scholar]
- 28.Anahtar MN, Gootenberg DB, Mitchell CM, Kwon DS. Cervicovaginal microbiota and reproductive health: the virtue of simplicity. Cell host & microbe. 2018. Feb 14;23(2):159–68. [DOI] [PubMed] [Google Scholar]
- 29.Klatt NR, Cheu R, Birse K, et al. Vaginal bacteria modify HIV tenofovir microbicide efficacy in African women. Science. 2017. Jun 2;356(6341):938–45. [DOI] [PubMed] [Google Scholar]
- 30.Zimmermann M, Zimmermann-Kogadeeva M, Wegmann R, Goodman AL. Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature. 2019. Jun;570(7762):462–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee CY, Cheu RK, Lemke MM, et al. Quantitative modeling predicts mechanistic links between pre-treatment microbiome composition and metronidazole efficacy in bacterial vaginosis. Nature communications. 2020. Dec 1;11(1):1–2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Onderdonk AB, Delaney ML, Fichorova RN. The human microbiome during bacterial vaginosis. Clinical microbiology reviews. 2016. Apr;29(2):223–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Morrison C, Fichorova RN, Mauck C, et al. Cervical inflammation and immunity associated with hormonal contraception, pregnancy, and HIV-1 seroconversion. JAIDS Journal of Acquired Immune Deficiency Syndromes. 2014. Jun 1;66(2):109–17. [DOI] [PubMed] [Google Scholar]
- 34.Caporaso JG, Lauber CL, Walters WA, et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. The ISME journal. 2012. Aug;6(8):1621–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Figure 1 – Dominant taxa at Baseline. Histogram breaking down the number of samples dominated by specific genera (A). Relative abundance of the dominant genus in each sample (B).
Supplemental Figure 2 – Longitudinal Lactobacillus levels. Levels of Lactobacillus tracked by patient over the course of the study.
Supplemental Figure 3 – immune Profile Relationship to 16 rRNA gene profile. Immune profiles generated from select soluble factors were assessed by Bray-Curtis dissimilarity and findings were visualized through PCoA; samples clustered by metataxonomic CT and facted by visit(A). Bray-Curtis analysis of CT classifications, facted by visit (B).
Supplemental Figure 4 – Baseline and follow up Beta diversity clustered by clearance profile. Bray-Curtis analysis of women from each clearance profile, faceted by visit. (A).
Supplemental Figure 5 – Alpha diversity metrics of richness, Shannon diversity, and inverse simpson diversity show positive correlation with Nugent score (A). Discordance between Nugent BV scores and and seqBV are visualized by the number of patients in each cervicotypes. Colors indicate the dominant taxa in each sample (B).
Supplemental Figure 6 – PCoA visualization of wUNIFRAC analysis of women by age, BMI, menstrual cycle day and stage, birth control strategy and self-reported race. (A).
Supplemental Table 1 – Patient characteristics. Adapted from Thurman et al.
Supplemental Table 2 – Soluble factors. Soluble factors from CVL were measured as described in Methods.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available in the Sequence Read Archive repository under ascension PRJNA691964.