

Abstract
Porcine circovirus 2 (PCV2) has been recognized as an immunosuppressive pathogen. However, the crosstalk between this virus and its host cells in related signaling pathways remains poorly understood. In this study, the expression profiles of 84 genes involved in transforming growth factor-beta (TGF-β) signaling pathway were probed in PCV2b-infected primary porcine alveolar macrophages (PAMs) by using an RT2 profiler PCR array system. The protein expression levels of cytokines involved in the TGF-β signaling pathway were determined with a RayBiotech fluorescent Quantibody® porcine cytokine array system. Results showed that 48, 30, and 42 genes were differentially expressed at 1, 24, and 48 h after infection, respectively. A large number of genes analyzed by a co-expression network and implicated in transcriptional regulation and apoptosis were differentially expressed in PCV2b-infected PAMs. Among these genes, TGF-β, interleukin-10, CCAAT/enhancer-binding protein beta (C/EBPB), growth arrest, and DNA-damage-inducible 45 beta (GADD45B), and BCL2 were upregulated. By contrast, SMAD family member 1 (smad1) and smad3 were downregulated. These results suggested that the TGF-β signaling pathway was repressed in PAMs at the early onset of PCV2 infection. The inhibited apoptosis was indicated by the upregulated C/EBPB, GADD45B, and BCL2, and by the downregulated smad1 and smad3, which possibly increased the duration of PCV2 replication-permissive conditions and caused a persistent infection. Our study may provide insights into the underlying antiviral functional changes in the immune system of PCV2-infected pigs.
Keywords: porcine circovirus 2, transforming growth factor-beta, mRNA, signaling pathway analysis
1. Introduction
Porcine circovirus 2 (PCV2), a non-enveloped, single-stranded circular DNA virus, is the primary causative pathogen of PCV2 systemic disease (PCV2-SD) and other PCV2 associated diseases (PCVADs). PCVADs are the most economically important diseases affecting the global swine industry [1,2]. A remarkable feature of PCV2-SD pigs is systemic immunosuppression. Pigs with PCV2-SD show severe depletion in lymphocytes in their lymphoid tissues and a remarkable decrease in T and B cells in their peripheral blood, suggesting that lymphoid depletion largely contributes to immunosuppression in PCV2-affected pigs [3,4,5]. The PCV2-induced decrease in the proliferation and lysis of lymphocytes [6,7] and apoptosis of primary lymphoid organs or precursor cells [8,9,10], are potential causes of lymphocyte depletion.
Cytokines, secreted by cells undergoing cellular stress, are essential for the development of the immune response. The differential expression of some cytokines, such as interleukin (IL)-1/8/10/12, and TNF-α, in PCV2-infected porcine monocytes and macrophages, have been reported [11,12,13,14,15]. IL-10 has been suggested to play an important role in PCV2-induced systemic immunosuppression. Transforming growth factor-beta (TGF-β) and its signaling pathway participates in cell growth, apoptosis, differentiation, migration, and metastasis in a context-dependent manner in various cell types [16,17]. Dysregulation of TGF-β signaling pathway has been implicated in numerous human diseases [18]. In addition, it is a pleiotropic cytokine and has regulatory activity on multiple types of immune cells. T cells are established as critical targets of TGF-β, which regulates T cell development, homeostasis, tolerance, and differentiation [19,20,21]. TGF-β plays its biological role primarily through the canonical Smads signaling pathway which has three isoforms that are involved in several developmental processes as TGF-βs, TβRs, and Smads [22]. Current reports indicated that the TGF-β and IL-10 play an important role in immunosuppression induced by porcine reproductive and respiratory syndrome virus (PRRSV) [23,24,25,26]. However, the involvement of TGF-β and its regulated downstream events in PCV2-induced systemic immunosuppression in PCV2-infected pigs is yet to be investigated. In this study, the early expression profile of TGF-β and its signaling pathway targets in porcine alveolar macrophage (PAM) inoculated with PCV2 were analyzed using a PCR array system. Our results will enhance our understanding of the role of TGF-β and its signaling pathway targets in the interaction of immune cells and PCV2.
2. Materials and Methods
2.1. Animals and Isolation of PAM Cells
Three one-month-old Landrace pigs free of PCV1, PCV2, swine fever virus, and PRRSV, as confirmed by PCR and serum antibody testing, were housed at the Pig Unit of the Laboratory Animal Center of Lanzhou Veterinary Research Institute.
Before the collection of alveolar fluid, piglets were euthanized with intravenous sodium pentobarbital overdose. PAMs were isolated from lungs according to previously described methods [27,28]. The isolated PAMs was adjusted to 1 × 107 cells/mL and cultured with a growth medium containing RPMI-1640 supplemented with 10% (v/v) fetal bovine serum (FBS, Gibco, Thermo Fisher Scientific Inc., Waltham, MA, USA), 100 U/mL penicillin, and 100 μg/mL streptomycin and incubated at 37 °C with 5% CO2.
2.2. Virus Preparation for Infection
PCV2b virus (ShDNY-PCV2, GenBank No. FJ948167, https://www.ncbi.nlm.nih.Gov/nuccore/FJ948167.1/, accessed on 11 August 2010) was propagated in porcine kidney 15 cell line (PK-15) as described previously [28]. PK-15 cells monolayers were inoculated at 60% confluence. After 24 h of incubation at 37 °C, the monolayer was treated with 200 mM D-glucosamine for 1 h. After further 72 h incubation, the cells were harvested and lysed three times by freeze-thawing. The genomic copies of the PCV2 in lysed cell mixture were determined by qPCR as previously described [29].
PAMs were infected with PCV2b in multiplicity of infection (MOI) 1 (1 viral DNA copy/cell) as described previously [28]. After infection, cells were cultured in a maintained medium RPMI-1640 supplemented with 2% FBS and incubated at 37 °C, and 5% CO2, for 1, 24, and 48 h respectively. The cells and supernatant were collected respectively at indicated time intervals. The cells were kept in RNAlater® RNA stabilization reagent kits (Qiagen, Hilden, Germany) to prevent RNA degradation and stored at −70 °C until use.
2.3. Detection of PCV2 in PAMs
PCV2 in PAMs after the infection was detected through immunofluorescence assay (IFA) [30]. The cells were fixed with 4% paraformaldehyde (Solarbio®, Beijing, China) and permeabilized with 0.1% TritonX-100 in PBS (Sigma, Burlington, MA, USA). PCV2 in PAMs was indicated by Quantum dots conjugated specific single domain antibody (QDs-psdAb) probe.
2.4. RNA Extraction and cDNA Preparation
Total RNA was extracted by using an RNA extraction kit (Qiagen, Hilden, Germany) according to the instruction. The quantity of RNA was determined with a microspectrophotometer (GE, Boston, MA, USA), and the RNA integrity was evaluated through 1.8% agarose gel electrophoresis. Up to 2 μg of total RNA was reverse transcribed to cDNA using an RT2 HT first strand kit (Qiagen, Hilden, Germany) according to the instruction. cDNA was stored at −20 °C for further quantitative PCR (qPCR) assay.
2.5. RT2 Profiler PCR Array Assay
TGF-β signaling pathway-related genes were detected by using RT2 Profiler PCR Array kits (PASS-235Z, Qiagen, Hilden, Germany) in the Mx3005p real-time PCR cycler (Agilent Technologies, Santa Clara, CA, USA) as described previously [28]. The mRNA expression levels of aim genes were analyzed using the 2−ΔΔCt methods by PCR Array system software version 3.5 (Qiagen, Hilden, Germany).
2.6. Analysis of TGF-β Expressions
The secreted TGF-β in the supernatant of PAMs post PCV2 infection was detected by Quantibody® porcine cytokine array system (RayBiotech, Guangzhou, China) as described previously [28].
2.7. Co-Expression Network Analysis
The expression data of 84 genes were subjected to co-expression network analysis. A gene co-expression network was built based on a gene co-expression measure by the R programming language. Pearson’s correlation coefficient was used for all possible gene pairs, and the corresponding p-value was calculated for each correlation. A total of 717 links pairs yielded p < 0.01. The network was visualized in Cytoscape 3.1.1 [31,32], and network topology was examined with the Network Analyzer tool in Cytoscape. MCODE, a Cytoscape plugin [33], was utilized to identify co-expression modules for the whole network.
2.8. Statistic Analysis
Statistical differences for PCR array and cytokine array data were evaluated by Student’s t-test. A p-value of less than 0.05 was considered significant.
3. Results
3.1. Confirmation of PCV2b Infection in PAMs
PCV2b-infected PAMs were monitored for the presence of PCV2 by IFA. IFA revealed the presence of virions of PCV2b in PAMs at 24 and 48 hpi compared with mock-infected control (Figure 1).
Figure 1.

PCV2 infection in PAMs. PCV2 in PAMs post-infection was detected by IFA based on the ODs-psdAb probe (red), the nucleus was stained with DAPI (blue).
3.2. MRNA Expression Profiles
To investigate the roles of TGF-β and its regulated downstream events in the interaction of PCV2 with host cells, PCV2b-infected PAMs were analyzed by using a PCR array kit. In this study gene expression differences between the control and treatment groups were considered significant using p < 0.05 and an absolute fold change ≥1.5 as a cut-off [34,35]. The fold change of differential gene expression is shown with the color scheme in the heatmap shown in Figure 2A. A total of 58 target genes involved in the TGF-β signaling pathway were differentially expressed (p < 0.05) in different stages of infection between the control and PCV2-infected groups (Table 1). At 1 hpi, 35 genes were upregulated, and 13 genes were downregulated. At 24 hpi, 21 genes were upregulated, and 9 genes were downregulated. At 48 hpi, 29 genes were upregulated, and 13 genes were downregulated.
Figure 2.

Gene expression patterns. Sixty-three target genes involved in the TGF-β signaling pathway were differentially expressed and were significantly different between PCV2-infected and control PAMs (A). Differentially expressed genes at each time point (B). Green: negative fold change level. Red: positive fold change level. Fold change was obtained by comparing the relative mRNA expression of target genes in PCV2-infected and uninfected control PAMs. Results are representative of three independent experiments and are presented as mean ± SE. ** represents p <0.01, * represents p < 0.05.
Table 1.
Differentially expressed TGF-β target genes (p < 0.05 and absolute fold change ≥1.5) between PCV2-infected and mock-infected control. Numbers in bold denote significant differential expression.
| Gene Symbol | Description | Fold Change | ||
|---|---|---|---|---|
| 1 hpi | 24 hpi | 48 hpi | ||
| Regulation of Transcription and Transcription Factor Activity | ||||
| C/EBPB | CCAAT/enhancer binding protein (C/EBP), beta | 4.96 | 2.87 | 2.55 |
| CREBBP | CREB binding protein | 8.75 | 4.33 | 3.66 |
| HMOX1 | Heme oxygenase (decycling) 1 | 7.53 | 4.49 | 3.50 |
| IL10 | Interleukin 10 | 5.97 | 4.69 | 4.68 |
| GADD45B | Growth arrest and DNA-damage-inducible, beta | 5.62 | 2.71 | 3.07 |
| NFKBIA | Nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha | 8.67 | 5.95 | 3.99 |
| RYBP | RING1 and YY1 binding protein | 3.42 | 1.59 | 1.54 |
| ID3 | Inhibitor of DNA binding 3, dominant negative helix-loop-helix protein | 1.28 | 6.84 | 28.03 |
| RUNX1 | Runt-related transcription factor 1 | 5.78 | 1.03 | 1.76 |
| MBD1 | Methyl-CpG binding domain protein 1 | 1.95 | 1.49 | 1.65 |
| SERPINE1 | Serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1), member 1 | 4.17 | –1.12 | 1.93 |
| ID2 | Inhibitor of DNA binding 2, dominant negative helix-loop-helix protein | 1.84 | –1.01 | 2.99 |
| ATF4 | Activating transcription factor 4 (tax-responsive enhancer element B67) | 2.08 | 1.26 | 1.15 |
| ATP1B1 | ATPase, Na+/K+ transporting, beta 1 polypeptide | 5.83 | 1.49 | −1.22 |
| BACH1 | BTB and CNC homology 1, basic leucine zipper transcription factor 1 | 5.48 | 1.06 | −1.04 |
| BHLHE40 | Basic helix-loop-helix family, member e40 | 2.76 | 1.32 | 1.43 |
| FOXO1 | Forkhead box O1 | 1.85 | 1.15 | 1.33 |
| PTK2B | PTK2B protein tyrosine kinase 2 beta | 1.79 | −1.01 | −1.01 |
| TXNIP | Thioredoxin interacting protein | 1.55 | −1.28 | 1.19 |
| VEGFA | Vascular endothelial growth factor-A | 12.49 | 1.52 | 1.91 |
| MAPK8 | Mitogen-activated protein kinase 8 | 1.54 | −1.01 | −1.31 |
| IFRD1 | Interferon-related developmental regulator 1 | 3.87 | 1.04 | −1.25 |
| LOC396709 | Retinoic acid receptor alpha | 2.89 | 1.15 | 1.17 |
| HSP90AA1 | 90-kDa heat shock protein | 1.50 | 1.36 | 1.15 |
| LOC100620797 | Focal adhesion kinase 1-like | 1.39 | 1.39 | 1.79 |
| CTNNB1 | Catenin (cadherin-associated protein), beta 1, 88 kDa | 1.43 | 1.67 | 1.02 |
| SP1 | Sp1 transcription factor | −1.01 | 1.52 | 1.41 |
| PTGS2 | Prostaglandin G/H synthase-2 | 21.61 | 2.25 | −5.99 |
| RGS4 | Regulator of G-protein signaling 4 | 2.39 | 7.21 | −2.49 |
| FURIN | Furin-like | 2.50 | −1.18 | −1.57 |
| JARID2 | Jumonji, AT rich interactive domain 2 | 1.63 | −1.12 | −1.74 |
| EMP1 | Epithelial membrane protein 1 | 3.35 | −1.52 | −3.65 |
| TNFSF10 | Tumor necrosis factor (ligand) superfamily, member 10 | −4.98 | 3.95 | 13.72 |
| MAPK14 | Mitogen-activated protein kinase 14 | −1.95 | 1.67 | 1.63 |
| STC2 | Stanniocalcin 2 | −2.06 | −1.57 | −1.52 |
| ME2 | Malic enzyme 2, NAD(+)-dependent, mitochondrial | −1.63 | −1.18 | −1.60 |
| GTF2I | General transcription factor IIi | −3.86 | −1.07 | 1.34 |
| PPARA | Peroxisome proliferator-activated receptor alpha | −3.69 | 1.41 | 1.39 |
| Regulation of programmed cell death and apoptosis | ||||
| TGFB2 | Transforming growth factor, beta 2 | 1.76 | 3.94 | 6.36 |
| S100A8 | S100 calcium binding protein A8 | 2.91 | 14.55 | 20.75 |
| SMAD6 | SMAD family member 6 | 2.10 | −1.01 | 1.44 |
| BCL2L1 | BCL2-like 1 | 1.16 | 1.68 | 1.59 |
| CDKN1B | Cyclin-dependent kinase inhibitor 1B (p27, Kip1) | −1.22 | 1.73 | 2.20 |
| FOS | FBJ murine osteosarcoma viral oncogene homolog | −2.63 | 2.37 | 18.83 |
| FN1 | Fibronectin 1 | 1.74 | −1.12 | 3.78 |
| MMP2 | Matrix metallopeptidase 2 (gelatinase A, 72kDa gelatinase, 72kDa type IV collagenase) | −1.96 | 1.23 | 2.88 |
| ACVR1 | Activin A receptor, type I | −1.15 | −1.58 | −1.82 |
| ACVRL1 | Activin A receptor type II-like 1 | −1.18 | −1.69 | −1.78 |
| MYC | V-myc myelocytomatosis viral oncogene homolog (avian) | −1.39 | −3.56 | −6.66 |
| HES1 | Hairy and enhancer of split 1, (Drosophila) | −1.33 | −2.15 | −3.09 |
| JUP | Junction plakoglobin | −1.30 | 1.00 | −3.28 |
| SMAD1 | SMAD family member 1 | −2.66 | −2.60 | 1.17 |
| SMAD3 | SMAD family member 3 | −2.12 | −4.28 | −1.08 |
| EPHB2 | EPH receptor B2 | −1.00 | −1.73 | −1.21 |
| No explicit function annotation | ||||
| CRYAB | Crystallin, alpha B | −1.25 | −2.25 | 1.56 |
| SREBF2 | Sterol regulatory element binding transcription factor 2 | −1.41 | −1.90 | −5.81 |
| ENG | Endoglin | 1.58 | 1.39 | 2.08 |
| BRF2 | Zinc finger protein 36, C3H type-like 2 | 1.33 | 1.21 | 1.93 |
In these differentially expressed genes, the immunosuppressive cytokines, TGF-β and IL-10 were upregulated significantly at 24 and 48 hpi. Smad family member 1 (smad1) and smad3, key mediators of the TGF-β signaling pathway, were downregulated significantly at 1 and 24 hpi. CCAAT/enhancer-binding protein beta (C/EBPB), growth arrest and DNA-damage-inducible 45 beta (GADD45B), and BCL2, which are associated with cell proliferation and apoptosis, were upregulated at different time points after PCV2 infection (Figure 2B).
3.3. Expression Analyses of TGF-β in PCV2b-Infected PAMs
PCR array results showed that TGF-β was upregulated significantly in PAMs post PCV2b infection. To further examine the expression in protein level, TGF-β in a culture supernatant of PAMs post PCV2b infection was detected by the Quantibody® porcine cytokine array kits. Results showed that the TGF-β was upregulated significantly in protein levels, which was consistent with the detected by PCR array (Figure 3).
Figure 3.

Expression of TGF-β at the protein level. TGF-β in the culture supernatants of control and PCV2-infected PAMs were measured by using porcine cytokine array kits. Data are expressed as pg/mL of three independent experiments. * represents p < 0.05.
3.4. Network Modularity
We constructed the co-expression network from the expression profiles of 84 TGF-β pathway-related genes. A total of 717 links were found in the network between the 84 genes, which were clustered to three highly interactive modules (Figure 4A) and showed high average clustering coefficients (Figure 4B). Functional analysis showed that the largest module had 32 enriched genes for the regulation of transcription. For these genes, 24 were upregulated and 5 were downregulated at 1 hpi, 7 were upregulated and 1 was downregulated at 24 hpi, and 10 were upregulated, and 5 were downregulated at 48 hpi. The second module with 14 genes was involved in the regulation of programmed cell death. For these genes, 3 were upregulated and 5 were downregulated at 1 hpi, 5 were upregulated and 5 were downregulated at 24 hpi, and 8 were upregulated, and 5 were downregulated at 48 hpi. The third module with seven genes had no explicit functional annotation. The gene expression levels of ID2 (2.99 fold at 48 hpi), ID3 (6.84 and 28.03 fold at 24 and 48 hpi), and SREBF2 (−1.9 and −5.81 fold at 24 and 48 hpi), which are implicated in the regulation of transcription, were significantly altered. TNFSF10, the apoptosis-related gene, was downregulated at 1 hpi but was upregulated at 24 and 48 hpi (Table 1).
Figure 4.
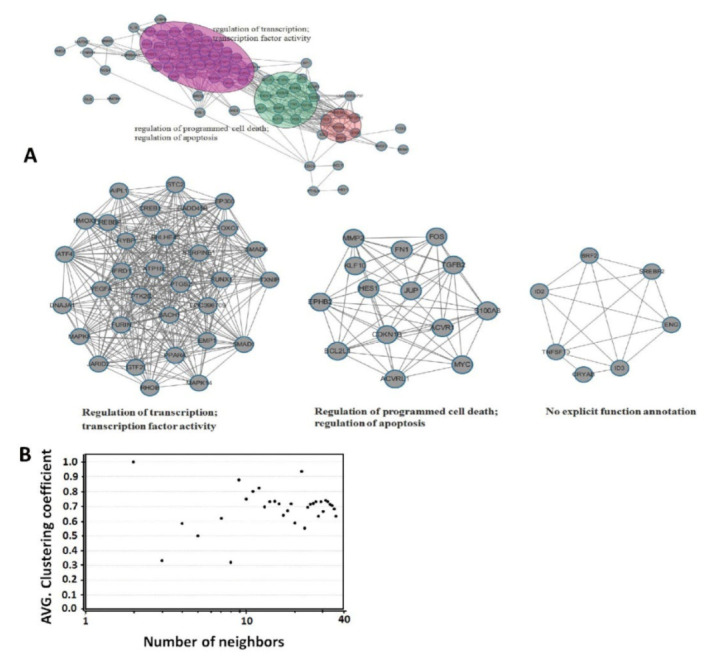
Co-expression network. A co-expression network from the expression profiles of 84 TGF-β signaling pathway-related genes was constructed. Interactive modules (A), Clustering coefficients (B).
4. Discussion
PCV2, one of the most important pathogens in domestic swine worldwide, is the causative agent of PCVAD, a widespread complex multi-factorial disease with recognized immunosuppressive characteristics. Vaccines against this virus display high levels of efficacy in controlling the infection [36,37], but the molecular mechanisms of immunosuppression in PCVAD remain largely unknown. PCV2 infection and replication in immune system cells are known as the potential mechanism for virus-induced immunosuppression. In addition, PCV2 causes immunosuppression by induction of apoptosis of immune system cells, and by acting as cytokines or cytokines inhibitors/enhancers [5,38]. Previous reports have shown that TGF-β is involved in apoptosis [39,40] and cell survival [41]. The present study aimed to analyze the expression profile of TGF-β and its signaling pathway target genes in the interaction of PCV2 with PAMs.
Monocyte/macrophage lineage cells play important roles in resisting the infection of pathogens. PCV2 has been shown to infect and persist in cells of the immune system, including PAMs [42,43,44]. PCV2 alone likely causes remarkable functional impairment in PAMs, including a reduction in phagocytosis and microbicidal capability [43]. Transcription analyses on PCV2-infected monocyte/macrophage lineage cells have shown that genes related to inflammation and apoptosis are differentially expressed [14]. In the present study, TGF-β signaling pathway target genes were analyzed in PCV2-infected PAMs by using a PCR array system. Meanwhile, the functions of 84 TGF-β signaling pathway target genes were explored by a co-expression network analysis. These genes were clustered into three highly interactive modules, which showed that their functions are in the regulation of transcription, programmed cell death, and apoptosis. The results of the PCR array showed that 58 of the 84 genes targeting the TGF-β signaling pathway were differentially expressed in PAM in different stages of post PCV2 infection. These results suggested that the TGF-β signaling pathway was possibly implicated in functional changes in PCV2-infected PAMs. Interestingly, 48 genes were differentially regulated at 1 hpi which only satisfy the adsorption and internalization for the virus [45]. Our results indicated that PAMs showed a strong reaction toward PCV2 during adsorption and internalization of the virus. Moreover, results suggested that the nucleic acid and structural protein of PCV2 may play a vital role in PAM’s reactivity. However, a comparative analysis of the differentially expressed genes in different infection stages revealed reduced host response of PAMs to PCV2-challenge. In particular, the genes involved in the regulation of transcription were reduced at 1 hpi (24 upregulated and 5 downregulated) to 48 hpi (10 upregulated and 5 downregulated). PAMs are the first line of defense cells to encounter PCV2 in the lungs and are involved in phagocytic clearance, inflammatory reactions, and tissue homeostasis. Thus, a decreased response to PCV2 may be a result of self-regulation of PAMs.
Whether gene expression leads to protein expression changes was determined by analyzing the differentially expressed genes TGF-β at the protein level with a Quantibody® array assay. The difference in TGF-β expression fitted well at mRNA and protein levels. Systemic immunosuppression is a remarkable feature of PCV2-infected pigs. TGF-β, which affects proliferation, differentiation, and apoptosis of immune cells through its signaling pathway, is also an immunosuppressive cytokine. In this study, TGF-β were upregulated significantly in PAM post-PCV2 challenge, implying that they may function in PCV2 pathogenesis.
TGF-β was significantly upregulated at mRNA and protein levels in PAM post-PCV2 infection. Smad1 and smad3, however, key mediators of the TGF-β signaling pathway for cell apoptosis [46,47,48] were downregulated. Moreover, BCL2, a hallmark protein participating in TGF-β-induced apoptosis and functioning pathophysiologically in apoptosis protection [49,50], was upregulated. In addition, C/EBPB and C/EBPB were upregulated. C/EBPB is involved in the differentiation of macrophages to functionally different subpopulations, and it is a key regulator for the survival of alveolar macrophages [51,52,53]. GADD45B, an anti-apoptotic factor, represses JNK-mediated apoptosis [54] and enhances the activation of C/EBPB. These two factors can function cooperatively in the regulation of cell differentiation [55]. Thus, the upregulation of BCL2, C/EBPB, and GADD45B, along with the downregulation of smad1 and smad3 may be a mechanism of apoptosis inhibition in PCV2-infected PAMs. These results suggested that even though TGF-β increased, PCV2 infection may have repressed TGF-β signaling pathway-induced apoptosis in PAMs. The inhibition of apoptosis will prolong the life of PAMs and may, therefore, contribute to PCV2 escaping host immunity and lengthen the duration of PCV2 replication-permissive conditions.
Global gene expression in PCV2-infected PAMs based on microarray analysis has been reported previously [14,56]. Similar to described for microarray analysis on PCV2-infected PAMs at an early stage (24 and 48 hpi), PCV2 seems to induce a large number of genes differential expression. Particularly, our research highlight the expression trend of EMP1, TNFSF10, S100A8, BCL2, and MYC genes involved in the regulation of transcription and apoptosis were the same as the microarray analysis [56]. However, the differentially expressed genes in this study, such as C/EBPB, CREBBP, IL-10, GADD45B, ID3, TGF-β (Table 1) were not shown in the differential expressed gene data of microarray analysis [14,56]. In addition, Mavrommatis et al. research [14] showed that PCV2 infection of PAMs (1 and 24 hpi) resulted in differential regulation of much fewer genes than described here or in Li et al. [56]. Whereas the differences between these results are not completely clear. The use of the different mock-infection, virus strains, and multiplicity of infection may be contributing to the differences between these results. Indeed, the virus strains and multiplicity of infection were different in these studies. Moreover, the treatment of cells and the reagents contaminated by LPS were potential reasons as well [14]. Furthermore, the different pig breeds (large white piglets, crossbred large white × landrace piglets were used in Li et al. [56] and Mavrommatis et al. [14] research respectively, landrace piglets was used here) and the potential microorganisms in PAMs (even though the specific pathogens are free) that may contribute to differential regulation of these genes.
5. Conclusions
PCR array analysis showed that 58 of the downstream genes of the TGF-β signaling pathway were differentially expressed in PAMs in different stages after PCV2 infection. Among these genes, IL-10 and TGF-β were upregulated significantly, suggesting their participation in PCV2 pathogenesis. However, the inhibited apoptosis in the PAMs indicated by the downregulated smad1 and smad3 and by the upregulated C/EBPB, GADD45B, and BCL2 may have partly prolonged PCV2 replication-permissive conditions and caused persistent infection in clinical cases. The observed expression profile of TGF-β signaling pathway-associated genes in PCV2-infected PAMs may provide a possible mechanism of the functional changes in the immune system of PCV2-affected pigs.
Author Contributions
S.Y. (Shunli Yang) developed the idea and drafted the project. M.U.Z.K. for his secretarial support in reviewing and editing the manuscript. B.L. for data analysis. S.Y. (Shuanghui Yin) helped with the isolation and culture of PAMs. M.H. and J.C. develop the results and discussion. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 31602085) and the Agricultural Science and Technology Innovation Program (CAAS-ASTIP-2014-LVRI-09). We are grateful for the help from all staff in the public instrument center of Lanzhou Veterinary Research Institute of the Chinese Academy of Agricultural Sciences.
Institutional Review Board Statement
All of the animals were handled in strict accordance with the Animal Ethics Procedures and Guidelines of the People’s Republic of China, and the current study was approved by the Animal Ethics Committee of Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences (No. LVRIAEC2020–035).
Informed Consent Statement
Not Applicable.
Data Availability Statement
Materials described in the manuscript are freely available to any scientist wishing to use them.
Conflicts of Interest
None of the authors of this paper has a financial or personal relationship with other people or organizations that could inappropriately influence the content of this paper.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Meng X.J. Spread like a wildfire--the omnipresence of porcine circovirus type 2 (PCV2) and its ever-expanding association with diseases in pigs. Virus Res. 2012;164:1–3. doi: 10.1016/j.virusres.2011.12.005. [DOI] [PubMed] [Google Scholar]
- 2.Ge X., Wang F., Guo X., Yang H. Porcine circovirus type 2 and its associated diseases in China. Virus Res. 2012;164:100–106. doi: 10.1016/j.virusres.2011.10.005. [DOI] [PubMed] [Google Scholar]
- 3.Segales J., Rose C., II, Domingo M. Pathological findings associated with naturally acquired porcine circovirus type 2 associated disease. Vet. Microbiol. 2004;98:137–149. doi: 10.1016/j.vetmic.2003.10.006. [DOI] [PubMed] [Google Scholar]
- 4.Darwich L., Segales J., Domingo M., Mateu E. Changes in CD4(+), CD8(+), CD4(+) CD8(+), and immunoglobulin M-positive peripheral blood mononuclear cells of postweaning multisystemic wasting syndrome-affected pigs and age-matched uninfected wasted and healthy pigs correlate with lesions and porcine circovirus type 2 load in lymphoid tissues. Clin. Diagn. Lab. Immunol. 2002;9:236–242. doi: 10.1128/CDLI.9.2.236-242.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Segales J., Domingo M., Chianini F., Majo N., Dominguez J., Darwich L., Mateu E. Immunosuppression in postweaning multisystemic wasting syndrome affected pigs. Vet. Microbiol. 2004;98:151–158. doi: 10.1016/j.vetmic.2003.10.007. [DOI] [PubMed] [Google Scholar]
- 6.Rodriguez-Carino C., Duffy C., Sanchez-Chardi A., McNeilly F., Allan G.M., Segales J. Porcine circovirus type 2 morphogenesis in a clone derived from the l35 lymphoblastoid cell line. J. Comp. Pathol. 2011;144:91–102. doi: 10.1016/j.jcpa.2010.07.001. [DOI] [PubMed] [Google Scholar]
- 7.Mandrioli L., Sarli G., Panarese S., Baldoni S., Marcato P.S. Apoptosis and proliferative activity in lymph node reaction in postweaning multisystemic wasting syndrome (PMWS) Vet. Immunol. Immunopathol. 2004;97:25–37. doi: 10.1016/j.vetimm.2003.08.017. [DOI] [PubMed] [Google Scholar]
- 8.Yu S., Opriessnig T., Kitikoon P., Nilubol D., Halbur P.G., Thacker E. Porcine circovirus type 2 (PCV2) distribution and replication in tissues and immune cells in early infected pigs. Vet. Immunol. Immunopathol. 2007;115:261–272. doi: 10.1016/j.vetimm.2006.11.006. [DOI] [PubMed] [Google Scholar]
- 9.Lin C.M., Jeng C.R., Hsiao S.H., Liu J.P., Chang C.C., Chiou M.T., Tsai Y.C., Chia M.Y., Pang V.F. Immunopathological characterization of porcine circovirus type 2 infection-associated follicular changes in inguinal lymph nodes using high-throughput tissue microarray. Vet. Microbiol. 2011;149:72–84. doi: 10.1016/j.vetmic.2010.10.018. [DOI] [PubMed] [Google Scholar]
- 10.Darwich L., Mateu E. Immunology of porcine circovirus type 2 (PCV2) Virus Res. 2012;164:61–67. doi: 10.1016/j.virusres.2011.12.003. [DOI] [PubMed] [Google Scholar]
- 11.Wikstrom F.H., Fossum C., Fuxler L., Kruse R., Lovgren T. Cytokine induction by immunostimulatory DNA in porcine PBMC is impaired by a hairpin forming sequence motif from the genome of Porcine Circovirus type 2 (PCV2) Vet. Immunol. Immunopathol. 2011;139:156–166. doi: 10.1016/j.vetimm.2010.09.010. [DOI] [PubMed] [Google Scholar]
- 12.Lee G., Han D., Song J.Y., Lee Y.S., Kang K.S., Yoon S. Genomic expression profiling in lymph nodes with lymphoid depletion from porcine circovirus 2-infected pigs. J. Gen. Virol. 2010;91:2585–2591. doi: 10.1099/vir.0.022608-0. [DOI] [PubMed] [Google Scholar]
- 13.Sun S.Q., Yin S.H., Guo H.C., Jin Y., Shang Y.J., Liu X.T. Genetic typing of classical swine fever virus isolates from China. Transbound. Emerg. Dis. 2013;60:370–375. doi: 10.1111/j.1865-1682.2012.01346.x. [DOI] [PubMed] [Google Scholar]
- 14.Mavrommatis B., Offord V., Patterson R., Watson M., Kanellos T., Steinbach F., Grierson S., Werling D. Global gene expression profiling of myeloid immune cell subsets in response to in vitro challenge with porcine circovirus 2b. PLoS ONE. 2014;9:e91081. doi: 10.1371/journal.pone.0091081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Qin Y., Li H., Qiao J. TLR2/MyD88/NF-kappaB signalling pathway regulates IL-8 production in porcine alveolar macrophages infected with porcine circovirus 2. J. Gen. Virol. 2016;97:445–452. doi: 10.1099/jgv.0.000345. [DOI] [PubMed] [Google Scholar]
- 16.Schuster N., Krieglstein K. Mechanisms of TGF-beta-mediated apoptosis. Cell Tissue Res. 2002;307:1–14. doi: 10.1007/s00441-001-0479-6. [DOI] [PubMed] [Google Scholar]
- 17.Lee H.J. The Role of Tripartite Motif Family Proteins in TGF-beta Signaling Pathway and Cancer. J. Cancer Prev. 2018;23:162–169. doi: 10.15430/JCP.2018.23.4.162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Li C., Ebert P.J., Li Q.J. T cell receptor (TCR) and transforming growth factor beta (TGF-beta) signaling converge on DNA (cytosine-5)-methyltransferase to control forkhead box protein 3 (foxp3) locus methylation and inducible regulatory T cell differentiation. J. Biol. Chem. 2013;288:19127–19139. doi: 10.1074/jbc.M113.453357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Oh S.A., Li M.O. TGF-beta: Guardian of T cell function. J. Immunol. 2013;191:3973–3979. doi: 10.4049/jimmunol.1301843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Li M.O., Sanjabi S., Flavell R.A. Transforming growth factor-beta controls development, homeostasis, and tolerance of T cells by regulatory T cell-dependent and -independent mechanisms. Immunity. 2006;25:455–471. doi: 10.1016/j.immuni.2006.07.011. [DOI] [PubMed] [Google Scholar]
- 21.Li M.O., Wan Y.Y., Flavell R.A. T cell-produced transforming growth factor-beta1 controls T cell tolerance and regulates Th1- and Th17-cell differentiation. Immunity. 2007;26:579–591. doi: 10.1016/j.immuni.2007.03.014. [DOI] [PubMed] [Google Scholar]
- 22.Derynck R., Zhang Y.E. Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature. 2003;425:577–584. doi: 10.1038/nature02006. [DOI] [PubMed] [Google Scholar]
- 23.Dwivedi V., Manickam C., Patterson R., Dodson K., Murtaugh M., Torrelles J.B., Schlesinger L.S., Renukaradhya G.J. Cross-protective immunity to porcine reproductive and respiratory syndrome virus by intranasal delivery of a live virus vaccine with a potent adjuvant. Vaccine. 2011;29:4058–4066. doi: 10.1016/j.vaccine.2011.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.LeRoith T., Hammond S., Todd S.M., Ni Y., Cecere T., Pelzer K.D. A modified live PRRSV vaccine and the pathogenic parent strain induce regulatory T cells in pigs naturally infected with Mycoplasma hyopneumoniae. Vet. Immunol. Immunopathol. 2011;140:312–316. doi: 10.1016/j.vetimm.2010.12.011. [DOI] [PubMed] [Google Scholar]
- 25.Renukaradhya G.J., Alekseev K., Jung K., Fang Y., Saif L.J. Porcine Reproductive and Respiratory Syndrome Virus-Induced Immunosuppression Exacerbates the Inflammatory Response to Porcine Respiratory Coronavirus in Pigs. Viral. Immunol. 2010;23:457–466. doi: 10.1089/vim.2010.0051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Renukaradhya G.J., Dwivedi V., Manickam C., Binjawadagi B., Benfield D. Mucosal vaccines to prevent porcine reproductive and respiratory syndrome: A new perspective. Anim. Health Res. Rev. 2012;13:21–37. doi: 10.1017/S1466252312000023. [DOI] [PubMed] [Google Scholar]
- 27.Olvera A., Ballester M., Nofrarias M., Sibila M., Aragon V. Differences in phagocytosis susceptibility in Haemophilus parasuis strains. Vet. Res. 2009;40:24. doi: 10.1051/vetres/2009007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yang S., Liu B., Yin S., Shang Y., Zhang X., Khan M.U.Z., Liu X., Cai J. Porcine Circovirus Type 2 Induces Single Immunoglobulin Interleukin-1 Related Receptor (SIGIRR) Downregulation to Promote Interleukin-1beta Upregulation in Porcine Alveolar Macrophage. Viruses. 2019;11:1021. doi: 10.3390/v11111021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.McIntosh K.A., Tumber A., Harding J.C., Krakowka S., Ellis J.A., Hill J.E. Development and validation of a SYBR green real-time PCR for the quantification of porcine circovirus type 2 in serum, buffy coat, feces, and multiple tissues. Vet. Microbiol. 2009;133:23–33. doi: 10.1016/j.vetmic.2008.06.010. [DOI] [PubMed] [Google Scholar]
- 30.Yang S.L., Shang Y.J., Wang D., Yin S.H., Cai J.P., Liu X.T. Diagnosis of porcine circovirus type 2 infection with a combination of immunomagnetic beads, single-domain antibody, and fluorescent quantum dot probes. Arch. Virol. 2015;160:2325–2334. doi: 10.1007/s00705-015-2508-x. [DOI] [PubMed] [Google Scholar]
- 31.Shannon P., Markiel A., Ozier O., Baliga N.S., Wang J.T., Ramage D., Amin N., Schwikowski B., Ideker T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13:2498–2504. doi: 10.1101/gr.1239303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Su G., Morris J.H., Demchak B., Bader G.D. Biological network exploration with cytoscape 3. Curr. Protoc. Bioinform. 2014;47:1–24. doi: 10.1002/0471250953.bi0813s47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bader G.D., Hogue C.W. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003;4:2. doi: 10.1186/1471-2105-4-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wu D., Lee Y.G., Liu H.C., Yuan R.Y., Chiou H.Y., Hung C.H., Hu C.J. Identification of TLR downstream pathways in stroke patients. Clin. Biochem. 2013;46:1058–1064. doi: 10.1016/j.clinbiochem.2013.05.059. [DOI] [PubMed] [Google Scholar]
- 35.Trager U., Andre R., Lahiri N., Magnusson-Lind A., Weiss A., Grueninger S., McKinnon C., Sirinathsinghji E., Kahlon S., Pfister E.L., et al. HTT-lowering reverses Huntington’s disease immune dysfunction caused by NFkappaB pathway dysregulation. Brain A J. Neurol. 2014;137:819–833. doi: 10.1093/brain/awt355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Young M.G., Cunningham G.L., Sanford S.E. Circovirus vaccination in pigs with subclinical porcine circovirus type 2 infection complicated by ileitis. J. Swine Health Prod. 2011;19:175–180. [Google Scholar]
- 37.Beach N.M., Meng X.J. Efficacy and future prospects of commercially available and experimental vaccines against porcine circovirus type 2 (PCV2) Virus Res. 2012;164:33–42. doi: 10.1016/j.virusres.2011.09.041. [DOI] [PubMed] [Google Scholar]
- 38.Drew T.W. A review of evidence for immunosuppression due to porcine reproductive and respiratory syndrome virus. Vet Res. 2000;31:27–39. doi: 10.1051/vetres:2000106. [DOI] [PubMed] [Google Scholar]
- 39.Chang C.J., Liao C.H., Wang F.H., Lin C.M. Transforming growth factor-beta induces apoptosis in antigen-specific CD4+ T cells prepared for adoptive immunotherapy. Immunol. Lett. 2003;86:37–43. doi: 10.1016/S0165-2478(02)00307-3. [DOI] [PubMed] [Google Scholar]
- 40.Heldin C.H., Landstrom M., Moustakas A. Mechanism of TGF-beta signaling to growth arrest, apoptosis, and epithelial-mesenchymal transition. Curr. Opin. Cell Biol. 2009;21:166–176. doi: 10.1016/j.ceb.2009.01.021. [DOI] [PubMed] [Google Scholar]
- 41.Sanchez-Capelo A. Dual role for TGF-beta1 in apoptosis. Cytokine Growth Factor Rev. 2005;16:15–34. doi: 10.1016/j.cytogfr.2004.11.002. [DOI] [PubMed] [Google Scholar]
- 42.Gilpin D.F., McCullough K., Meehan B.M., McNeilly F., McNair I., Stevenson L.S., Foster J.C., Ellis J.A., Krakowka S., Adair B.M., et al. In vitro studies on the infection and replication of porcine circovirus type 2 in cells of the porcine immune system. Vet. Immunol. Immunopathol. 2003;94:149–161. doi: 10.1016/S0165-2427(03)00087-4. [DOI] [PubMed] [Google Scholar]
- 43.Chang H.W., Jeng C.R., Lin T.L., Liu J.J., Chiou M.T., Tsai Y.C., Chia M.Y., Jan T.R., Pang V.F. Immunopathological effects of porcine circovirus type 2 (PCV2) on swine alveolar macrophages by in vitro inoculation. Vet. Immunol. Immunopathol. 2006;110:207–219. doi: 10.1016/j.vetimm.2005.09.016. [DOI] [PubMed] [Google Scholar]
- 44.Chang H.W., Jeng C.R., Liu J.J., Lin T.L., Chang C.C., Chia M.Y., Tsai Y.C., Pang V.F. Reduction of porcine reproductive and respiratory syndrome virus (PRRSV) infection in swine alveolar macrophages by porcine circovirus 2 (PCV2)-induced interferon-alpha. Vet. Microbiol. 2005;108:167–177. doi: 10.1016/j.vetmic.2005.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Conery A.R., Cao Y., Thompson E.A., Townsend C.M., Jr., Ko T.C., Luo K. Akt interacts directly with Smad3 to regulate the sensitivity to TGF-beta induced apoptosis. Nat. Cell Biol. 2004;6:366–372. doi: 10.1038/ncb1117. [DOI] [PubMed] [Google Scholar]
- 46.Akhurst R.J., Hata A. Targeting the TGFbeta signalling pathway in disease. Nat. Rev. Drug Discov. 2012;11:790–811. doi: 10.1038/nrd3810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Heldin C.H., Miyazono K., ten Dijke P. TGF-beta signalling from cell membrane to nucleus through SMAD proteins. Nature. 1997;390:465–471. doi: 10.1038/37284. [DOI] [PubMed] [Google Scholar]
- 48.Abdollah S., MaciasSilva M., Tsukazaki T., Hayashi H., Attisano L., Wrana J.L. T beta RI phosphorylation of Smad2 on Ser(465) and Ser(467) is required for Smad2-Smad4 complex formation and signaling. J. Biol. Chem. 1997;272:27678–27685. doi: 10.1074/jbc.272.44.27678. [DOI] [PubMed] [Google Scholar]
- 49.Letai A.G. Diagnosing and exploiting cancer’s addiction to blocks in apoptosis. Nat. Rev. Cancer. 2008;8:121–132. doi: 10.1038/nrc2297. [DOI] [PubMed] [Google Scholar]
- 50.Cory S., Huang D.C.S., Adams J.M. The Bcl-2 family: Roles in cell survival and oncogenesis. Oncogene. 2003;22:8590–8607. doi: 10.1038/sj.onc.1207102. [DOI] [PubMed] [Google Scholar]
- 51.Cain D.W., O’Koren E.G., Kan M.J., Womble M., Sempowski G.D., Hopper K., Gunn M.D., Kelsoe G. Identification of a tissue-specific, C/EBPbeta-dependent pathway of differentiation for murine peritoneal macrophages. J. Immunol. 2013;191:4665–4675. doi: 10.4049/jimmunol.1300581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ruffell D., Mourkioti F., Gambardella A., Kirstetter P., Lopez R.G., Rosenthal N., Nerlov C. A CREB-C/EBPbeta cascade induces M2 macrophage-specific gene expression and promotes muscle injury repair. Proc. Natl. Acad. Sci. USA. 2009;106:17475–17480. doi: 10.1073/pnas.0908641106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Tamura A., Hirai H., Yokota A., Sato A., Shoji T., Kashiwagi T., Iwasa M., Fujishiro A., Miura Y., Maekawa T. Accelerated apoptosis of peripheral blood monocytes in Cebpb-deficient mice. Biochem. Biophys. Res. Commun. 2015;464:654–658. doi: 10.1016/j.bbrc.2015.07.045. [DOI] [PubMed] [Google Scholar]
- 54.Papa S., Zazzeroni F., Bubici C., Jayawardena S., Alvarez K., Matsuda S., Nguyen D.U., Pham C.G., Nelsbach A.H., Melis T., et al. Gadd45 beta mediates the NF-kappa B suppression of JNK signalling by targeting MKK7/JNKK2. Nat. Cell Biol. 2004;6:146–153. doi: 10.1038/ncb1093. [DOI] [PubMed] [Google Scholar]
- 55.Tsuchimochi K., Otero M., Dragomir C.L., Plumb D.A., Zerbini L.F., Libermann T.A., Marcu K.B., Komiya S., Ijiri K., Goldring M.B. GADD45beta enhances Col10a1 transcription via the MTK1/MKK3/6/p38 axis and activation of C/EBPbeta-TAD4 in terminally differentiating chondrocytes. J. Biol. Chem. 2010;285:8395–8407. doi: 10.1074/jbc.M109.038638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Li W., Liu S., Wang Y., Deng F., Yan W., Yang K., Chen H., He Q., Charreyre C., Audoneet J.C. Transcription analysis of the porcine alveolar macrophage response to porcine circovirus type 2. BMC Genom. 2013;14:353. doi: 10.1186/1471-2164-14-353. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Materials described in the manuscript are freely available to any scientist wishing to use them.