
Figure 12.
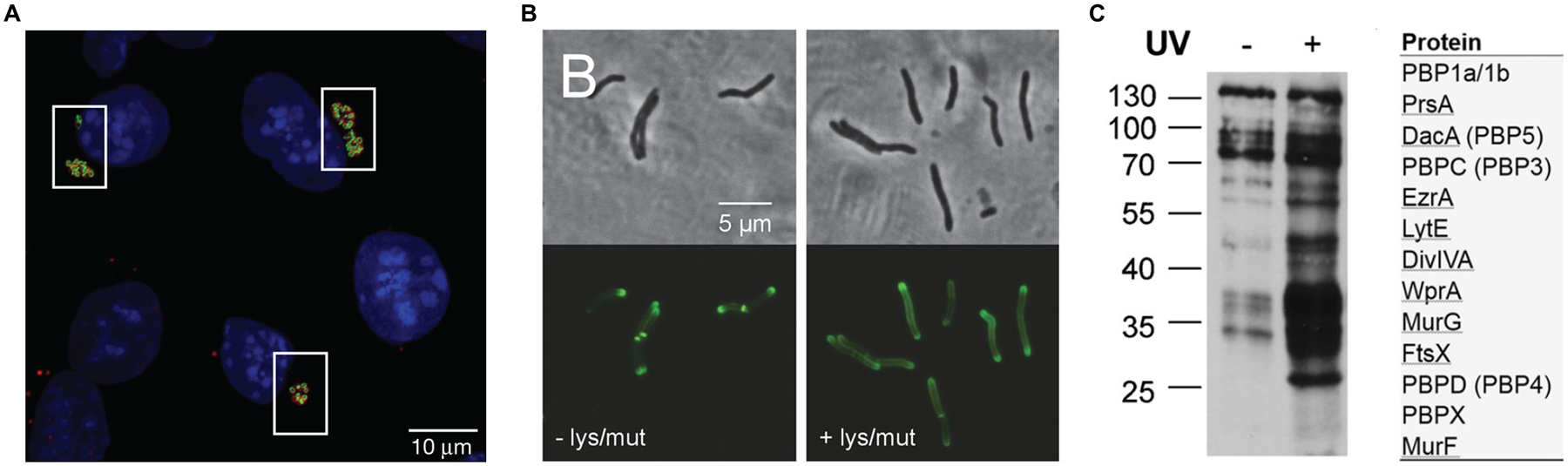
Imaging and proteomics applications of D-amino acid dipeptide reporters for PG. (A) First direct detection of PG in Chlamydia. Mouse fibroblasts were infected with C. trachomatis in the presence of AlkDADA (28), fixed, subjected to CuAAC with azido-488, and imaged. Blue, DAPI nuclear staining of host cells; green, C. trachomatis PG labeling; red, C. trachomatis major outer membrane protein (MOMP) staining. Reproduced with permission from ref 83. Copyright 2014 Springer Nature. (B) Mycobacterial sidewall synthesis of PG in response to cell wall damage. M. smegmatis was incubated in the absence (−) or presence (+) of PG-damaging enzymes lysozyme (lys) and mutanolysin (mut), then treated with AlkDADA (28), fixed, subjected to CuAAC with azido-488, and imaged. While polar labeling is predominant in the -lys/mut condition, enhanced peripheral labeling is visually evident in the +lys/mut condition, and was quantified in ref 110. Reproduced from ref 110. (C) Identification of Lipid II-interacting proteins in B. subtilis. Bacteria were incubated in photo-cross-linking probe x-DADA-Alk (32), exposed to UV irradiation, subjected to CuAAC with azido-biotin, and then biotinylated proteins were either detected by Western blot (left) or avidin-enriched, trypsinized, and analyzed by LC-MS/MS (right). Several PBPs and other PG-related proteins were identified. Reproduced with permission from ref 84. Copyright 2016 Wiley-VCH.