
Fig. 3. Mammalian sirtuins and metabolic reprogramming.
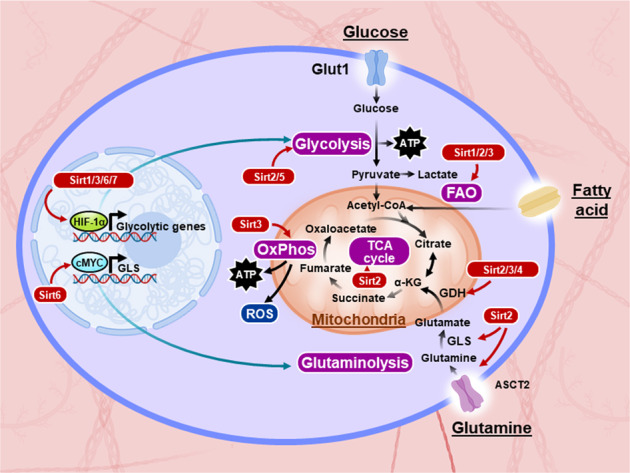
The entry of glucose into the cell is mediated by the glucose transporter Glut1. Glucose is then metabolized to pyruvate, which enters the mitochondrial tricarboxylic acid (TCA) cycle and generates reducing equivalents for ATP production via oxidative phosphorylation (OxPhos). During aerobic glycolysis, pyruvate is fermented into lactate in the cytoplasm despite the availability of oxygen for complete glucose oxidation; this process is called the Warburg effect. Glutamine enters cells using alanine, serine, cysteine-preferring transporter 2 (ASCT2), and it is converted into glutamate by glutaminase (GLS) and into α-ketoglutarate (α-KG) by glutamate dehydrogenase (GDH), which enters the TCA cycle for ATP production via OxPhos. Fatty acid molecules are catabolized into acetyl-CoA via fatty acid oxidation (FAO), and acetyl-CoA enters the TCA cycle for ATP production. Sirtuins are metabolic sensors that modulate a variety of metabolic pathways. Sirt1, Sirt3, and Sirt6 restrain the glycolytic pathway through hypoxia-inducible factor 1-alpha (HIF-1α) inhibition or direct effects. Sirt3 upregulates the OxPhos pathway by enhancing the activity of mitochondrial complexes I, II, and III and dampening reactive oxygen species (ROS) production. Sirt1 is also able to increase FAO by activating PPAR-α and PGC1, while Sirt3 upregulates FAO under caloric restriction conditions. Sirt2 deacetylates and inhibits the activity of many metabolic enzymes involved in glycolysis, glutaminolysis, the TCA cycle and FAO. Sirt3 and Sirt4 activate and inhibit glutaminolysis, respectively, by regulating GDH activity. Sirt6 impacts the glutaminolysis pathway by regulating c-Myc transcriptional activity. Sirt5 increases glycolysis by increasing the activity of the glycolytic enzyme GAPDH. Sirt7 can also repress HIF-1α and, therefore, may inhibit the transcription of glycolytic genes.