

Abstract
Chronic myeloid leukemia (CML) is a myeloproliferative disorder characterized by accumulation of immature cells in bone marrow and peripheral blood. Although successful results were obtained with tyrosine kinase inhibitors, several patients showed resistance. For this reason, the identification of new strategies and therapeutic biomarkers represents an attractive goal. The role of transient receptor potential (TRP) ion channels as possible drug targets has been elucidated in different types of cancer. Among natural compounds known to activate TRPs, cannabidiol (CBD) displays anticancer properties. By using FACS analysis, confocal microscopy, gene silencing, and cell growth assay, we demonstrated that CBD, through TRPV2, inhibits cell proliferation and cell cycle in CML cells. It promoted mitochondria dysfunction and mitophagy as shown by mitochondrial mass reduction and up‐regulation of several mitophagy markers. These effects were associated with changes in the expression of octamer‐binding transcription factor 4 and PU.1 markers regulated during cellular differentiation. Interestingly, a synergistic effect by combining CBD with the standard drug imatinib was found and imatinib‐resistant cells remain susceptible to CBD effects. Therefore, the targeting of TRPV2 by using CBD, through the activation of mitophagy and the reduction in stemness, could be a promising strategy to enhance conventional therapy and improve the prognosis of CML patients.
Keywords: cannabidiol, chronic myeloid leukemia, imatinib, mitophagy, TRPV2
Our data showed that cannabidiol, via TRPV2, induces cell proliferation blockage, mitophagy, and changes in the expression of differentiation markers in chronic myeloid leukemia cells. Cannabidiol acts synergistically with imatinib and it also reduces cell viability of imatinib‐resistant cells.
Abbreviations
- [Ca2+]i
Ca2+ influx
- ∆Ψm
mitochondrial transmembrane potential
- ATG16L1
autophagy protein 16 like 1
- ATG5‐ATG12
autophagy protein 12
- BAF
bafilomycin A
- CBD
cannabidiol
- CI
combination index
- CML
chronic myeloid leukemia
- COX IV
cytochrome c oxidase subunit 4
- DCFDA
2′,7′‐dichlorofluorescin diacetate
- H2O2
hydrogen peroxide
- JC‐1
5,5′,6,6′‐Tetrachloro‐1,1′,3,3′‐tetraethylbenzimidazolocarbocyanine iodide
- K562 IR
K562 imatininb‐resistant
- LC3
microtubule‐associated protein‐1 light chain 3
- MFI
mean fluorescence intensity
- OCT4
octamer‐binding transcription factor 4
- PI
propidium iodide
- pink1
PTEN induced putative kinase 1
- qRT‐PCR
quantitative real‐time PCR
- ROS
reactive oxygen species
- TKIs
tyrosine kinase inhibitors
- TRP
transient receptor potential
1. INTRODUCTION
Chronic myeloid leukemia (CML) is a hematologic malignancy which progresses in stages termed chronic phase, accelerated phase, and blast. 1 In addition to allogeneic stem cell transplantation or interferon administration, TKIs are the current approved therapy. However, the resistance to TKIs represents a limit that leads to therapy discontinuity associated with reduced survival of patients. 1 , 2 Consequently, the development of new therapeutic strategies and the identification of new therapeutical targets is getting attention.
In this regard, the role of ion channels belonging to the TRP family as possible targets, being implicated in cancer growth, metastasis, and chemoresistance, 3 has been established. The clear association with cancers and the possibility to control their functions by several modulators make them attractive targets for pharmacological intervention. 4
Few data are present about the involvement of TRP channels in hematological malignancies. The expression of several members of the TRP family in different lymphoid and myeloid leukemia has been demonstrated. 5 , 6 In fact, it has been shown that TRPV5 and 6‐induced [Ca2+]i stimulates cell cycle arrest in myeloid leukemia cell lines. 7 The inhibition of TRPM7 reduces cell proliferation and promotes erythroid differentiation in K562 CML cells. 8 Moreover, in this cell line, TRPV2 channel shows altered gene expression profile that is in part involved in the growth of blast cells. 9
Several natural compounds are known to modulate the TRP activities 10 and among them cannabidiol (CBD), a no‐psychoactive phytochemical extracted from Cannabis sp., has recently attracted consideration for its ability to act as an agonist for TRPV1 and TRPV2, while it has no affinity for the endocannabinoid CB1 and CB2 receptors. 11
CBD has been shown to reduce cell viability in immortalized human T lymphocyte cells and acute leukemic T cells 12 ; however, findings about the anticancer properties of CBD in CML cells are still missing.
Thus, this study investigated the effects induced by CBD treatment via activating TRPV2 in CML cell lines. In particular, given that in hematological malignancies the targeting of mitochondria increases the therapy efficacy, 13 we focused the attention on the mitophagy and the modulation of markers involved in the differentiation of CML cells. Finally, the combination of CBD with imatinib was also studied.
2. MATERIALS AND METHODS
2.1. Cell lines
Human K562, KU812 and MOLM‐6 CML cell lines, from DSMZ–German Collection of Microorganisms and Cell Cultures GmgH, were maintained in RPMI1640 medium (Euroclone) supplemented with 10% FBS (Euroclone), 2 mM l‐glutamine, 100 IU/mL of penicillin, and 100 μg/mL of streptomycin. K562 IR was developed by exposing cells to a concentration of 1 µM imatinib for 10 days. 14
Human acute leukemia THP‐1 cell line, provided by IFOM (Rome, Italy), was cultured in RPMI‐1640 Medium supplemented as above described and with 2‐mercaptoethanol (0.05 mM).
Cell lines were cultured at 37°C, 5% CO2 and 95% of humidity.
Blood of healthy donors, provided by Macerata Hospital (Italy) after authorization, was used to isolate PBMCs/red blood cells by Ficoll paque (Cederlane) and to obtain an enrichment of normal myeloid cells by a Rosette SepTM HLA Myeloid Cell Enrichment Kit (StemCell Technologies).
A CD38 Micro Bead Kit (Miltenyi Biotech Inc.) was used to isolate CD38+ common myeloid progenitors from human cord blood CD34+ progenitor cells (Sigma Aldrich). 15
2.2. Reagents
Capsazepine, PI, imatinib mesylate, ionomycin and DCFDA were from Sigma Aldrich. BAF, Fluo‐3 AM, JC‐1, and DAPI were from ThermoFisher Scientific. CBD was from ENECTA. Tranilast was from Bio‐Techne SRL. CBD, tranilast, capsazepine and BAF were dissolved in DMSO used as vehicle (maximum 0.05%, considered no toxic). 16 Abs used according to datasheet: anti‐LC3, anti‐COX IV, anti‐OCT4, anti‐ATG16L1, anti‐ATG5‐ATG12, anti‐pink1, anti‐parkin, anti‐caspase 3, anti‐GAPDH, anti‐optineurin and anti‐PU.1 were from Cell Signaling Technology (1:1000); rabbit anti‐human TRPV1 (1:1000) was from Invitrogen and goat anti‐human TRPV2 (1:50) was from Santa Cruz Biotechnology).
Secondary Abs used: HRP‐conjugated anti‐rabbit IgG (1:5000; Jackson ImmunoResearch Europe Ltd), HRP‐conjugated anti‐mouse IgG (1:2000; Cell Signaling Technology), PE‐conjugated goat anti‐rabbit Ab (1:40; BD Biosciences), FITC‐conjugated goat anti‐mouse Ab (1:40; BD Biosciences), Alexa Fluor‐594‐conjugated goat anti‐rabbit Ab (1:100; Cell Signaling Technology), and Alexa Fluor‐488‐conjugated donkey anti‐goat Ab (1:100; Invitrogen).
2.3. Quantitative real‐time PCR
Total RNA from CD34+CD38+ progenitors was extracted by SingleShot Cell Lysis Kit (BioRad) whereas the RNeasy Mini Kit (Qiagen) was used for all the other cells. cDNA was synthesized using the iScript Advanced cDNA Synthesis Kit (BioRad). qRT‐PCR reactions were performed with QuantiTect Primer Assays (Qiagen) for human TRPV1 (QT00046109), TRPV2 (QT00035987) and GAPDH (QT00079247), which was used as reference gene, using the iQ5 Multicolor Real‐Time PCR Detection System (BioRad). The PCR parameters were according to the primer datasheet. All samples were assayed in triplicate.
2.4. FACS analysis
Cells, fixed in paraformaldehyde 4% and permeabilized with cold methanol and permeabilization solution (1% FBS, 0.1% saponin, 0.1% sodium azide in PBS) were incubated with anti‐TRPV1, anti‐TRPV2, anti‐COX IV or anti‐OCT‐4 Abs. Cells were then incubated with fluorochrome‐conjugated secondary Abs and analyzed by FACS and CellQuest software (Beckton Dickinson).
2.5. Confocal microscope analysis
Chronic myeloid leukemia cells were fixed with 4% paraformaldehyde for 10 minutes and cytospin was performed. Cells were permeabilized using cold methanol and incubated with 5% of BSA and 0.1% of Tween‐20 in PBS for 1 hour at room temperature. After, cells were co‐labeled with anti‐TRPV1 Ab and anti‐TRPV2 Ab followed by fluorochrome‐conjugated secondary Abs. Nuclei were stained with DAPI. Slides were analyzed with C2 Plus confocal microscope (Nikon Instruments). Magnification = 100×.
Cells, treated or not with CBD at IC50 dose for 24 hours, were labelled with PU.1 Ab.
2.6. Intracellular calcium influx [Ca2+]i
1.5 × 106/mL CML cells were washed in calcium and magnesium free PBS supplemented with 4.5 g/L of glucose and then incubated with Fluo 3‐AM according to datasheet. After washing, cells, resuspended in PBS/glucose medium containing 2 mmol/L Ca2+, were stimulated with CBD (IC50 dose) or vehicle up to 3 minutes. Cells were also pretreated with tranilast before CBD. Ionomycin (5 µg/mL) treatment was used as positive control. Fluo 3‐AM fluorescence was measured by FACS and CellQuest Software.
2.7. Gene silencing
siTRPV2, siTRPV1 and siGLO nontargeting siRNA (used as control) FlexiTube siRNA were from Qiagen. CML cells were plated at 6 × 105/mL for 24 hours; the day after, cells were plated at the density of 4 × 105/mL and siTRPV2, siTRPV1 or siGLO (50 nM) were added, following the HiPerfect transfection reagent (Qiagen). Cells were harvested at 48 hours post‐transfection. Silencing efficiency was evaluated by qRT‐PCR and western blot. No differences were observed comparing siGLO transfected with untransfected cells.
2.8. Cell viability assay
2 × 105 CML cells/mL were plated in a 12‐well plate. The day after, CBD (10‐75 μM) or vehicle was added for 24 hours. Then, cells were stained with trypan blue and counted by using the TC20 automated cell counter (BioRad). Three replicates were used for each treatment. IC50 values were calculated using GraphPad Prism® 5.0 (GraphPad Software).
Chronic myeloid leukemia cells were also pretreated with tranilast (10 μM) or with capsazepine (10 μM) for 1 hour before the addition of CBD for 24 hours. Cell counting was performed in siTRPV2, siTRPV1 or siGLO CML cells treated with CBD (IC50) or vehicle. CBD was used in combination with imatinib mesylate for 24 hours. Synergistic activity of the CBD/imatinib combination was determined by the isobologram analysis and combination index (CI) methods (CompuSyn Software, ComboSyn, Inc.). The CI was used to express synergism (CI < 1), additivity (CI = 1) or antagonism (CI > 1).
Cell growth was measured in: cells pretreated with BAF (25 nM) for 1 hour and then treated with CBD (IC50 dose) for 24 hours, in K562 IR or K562 treated with imatinib (0.01‐10 μM) and in K562 IR cells treated for 24 hours with different doses of CBD.
2.9. Western blot analysis
Lysate from CML cells, treated or not with CBD at IC50 dose, was extracted by using lysis‐buffer (10 mM Tris, 100 mM sodium chloride, 1 mM EDTA, 1 mM EGTA, 1 mM sodium fluoride, 20 mM sodium pyrophosphate, 2 mM sodium orthovanadate, 1% Triton X‐100, 10% glycerol, 0.1% SDS, 0.5% deoxycholate, and 1 mM PMSF) containing protease‐inhibitor cocktail (EuroClone). Proteins were separated on 8%‐14% SDS polyacrylamide gels and transferred. Blocking was with 5% low‐fat dry milk or 5% BSA in PBS 0.1% Tween 20. Membranes were incubated with anti‐TRPV1, anti‐TRPV2, anti‐LC3, anti‐ATG16L1, anti‐ATG12, anti‐PINK1, anti‐optineurin, anti‐PU.1, anti‐parkin or anti‐GAPDH Abs followed by corresponding HRP‐conjugated secondary Abs. Analysis was performed by LiteAblot PLUS kit, Chemidoc and the Quantity One software (BioRad).
Lysates from CML cells treated with CBD at IC50 dose in combination with BAF for 12 hours or from siTRPV2 and siGLO (control) CML cells treated or not with CBD were incubated with anti‐LC3. Lysates from KU812 cells treated or not with CBD 15 µM for 24 hours were used for caspase‐3 analysis.
2.10. Cell cycle analysis
Chronic myeloid leukemia cells treated or not with CBD at IC50 dose for 24 hours were fixed in cold 70% ethanol, treated for 30 minutes at 37°C with 100 μg/mL ribonuclease A solution, stained for 30 minutes at room temperature with PI 20 μg/mL, and analyzed by FACS. CML cells were also pre‐treated with tranilast (10 μM) for 1 hour before the addition of CBD at dose IC50 for 24 hours.
2.11. BrdU assay
Cell proliferation was evaluated using BrdU staining kit (ThermoFisher Scientific) according to the company protocol. Cells were then analyzed by FACS and CellQuest software.
2.12. Cell death analysis
Cells, treated or not with CBD at IC50 dose for 24 hours, were incubated with 2 μg/mL PI for 30 minutes at 37°C. After washing, fluorescence was analyzed by FACS and CellQuest software. PBMCs or red blood cells were treated with CBD 25 µM or not for 24 hours and then Annexin V and/or PI staining was performed according to the datasheet (Enzo Life Science). As positive control, PBMCs and red blood cells were treated with hydrogen peroxide (H2O2). 17 , 18
2.13. Reactive oxygen species production
Oxidative stress levels in CML cells after treatment with CBD (IC50) was assessed by staining with 20 μM DCFDA for 20 minutes. After washing, the fluorescence was assayed using FACS and CellQuest software. Moreover, oxidative stress was evaluated in CBD (25 µM)‐treated red blood cells. As positive control, red blood cells were treated with H2O2. 18
2.14. Mitochondrial transmembrane potential (∆Ψm) and mitochondrial integrity
∆Ψm was evaluated by JC‐I staining according to the company protocol in CML cells, treated with CBD at IC50 dose or not for 8, 12 and 24 hours. Samples were then analyzed by FACS and CellQuest software. The drop of ∆Ψm was also evaluated in siGLO and siTRPV2 cells treated with vehicle or with CBD (IC50) for 12 hours.
Moreover, JC‐1 was used to evaluate ∆Ψm in healthy PBMCs treated or not with CBD (25 µM). The mitochondrial integrity was also investigated by labeling cells with Mitobright (Dojindo Molecular Technologies) according to instructions.
2.15. Mitophagy assay
Mitophagy in CML cells, treated with CBD at IC50 dose or with vehicle for 24 hours, was detected using the Mtphagy detection kit® (Dojindo Molecular Technologies). CML cells (105 cells/well) were labeled with 100 nM Mtphagy Dye for 15 minutes before the addition of CBD for 24 hours. Then cells were analyzed by FACS and CellQuest software. Confocal microscopy was also used. CML cells, stained with Mtphagy dye and treated with CBD as above described, were finally labelled with 1 µM Lyso Dye. After washes, cytospin was performed. Nuclei were counteracted with DAPI before the analysis with C2 Plus confocal laser scanning microscope by using 100× magnification.
2.16. Colony formation assay
The clonogenic activity of CML cells was measured with the Human Colony‐Forming Unit (CFU) Assay using MethoCult kit (StemCell Technologies). CML cells were treated for 24 hours with CBD (IC50 dose) or vehicle and then 2000 cells/well were plated according to the datasheet. After 10 days, colonies were visualized and counted by inverted bright field microscope. Each sample was assayed in triplicate.
2.17. Statistical analysis
The statistical significance was determined by Student's t test and by ANOVA with Bonferroni's post‐test. No statistically significant differences were found between untransfected and siGLO‐transfected CML cells, untreated with vehicle‐treated cells or vehicle‐treated cells at different times (data not shown). Given that no differences were found between vehicle‐treated cells at different times, for simplicity, as control, in time course analysis, vehicle‐treated cells for 24 h were shown.
3. RESULTS
3.1. CBD, by activating TRPV2, affects CML cell viability
We evaluated the expression of TRPV1 and TRPV2 in K562, KU812, MOLM‐6 CML cell lines, CD34+CD38+ common myeloid progenitors, normal myeloid cells, PBMCs and THP‐1 cells used as control. 9 , 19 TRPV1 and TRPV2 are expressed at mRNA level in all analyzed cells with TRPV2 that is expressed at higher levels in K562 and KU812 cells respect to THP‐1 (Figure 1A). The expression was also confirmed at protein levels by immunoblotting and cytofluorimetric analysis (Figure 1B,C). Moreover, confocal microscopy showed that TRPV1 and TRPV2 partially colocalize (Figure 1D). Given that CBD is able to activate both TRPV1 and TRPV2, 20 , 21 CML cells were treated with CBD (10‐75 μM) for 24 hours and analyzed by cell viability assay. CBD induced a significant decrease of cell viability with an IC50 of 20 μM (K562), 15 μM (KU812), and 25 μM (MOLM‐6) (Figure 1E). The IC50 doses of CBD were used for the subsequent experiments.
FIGURE 1.
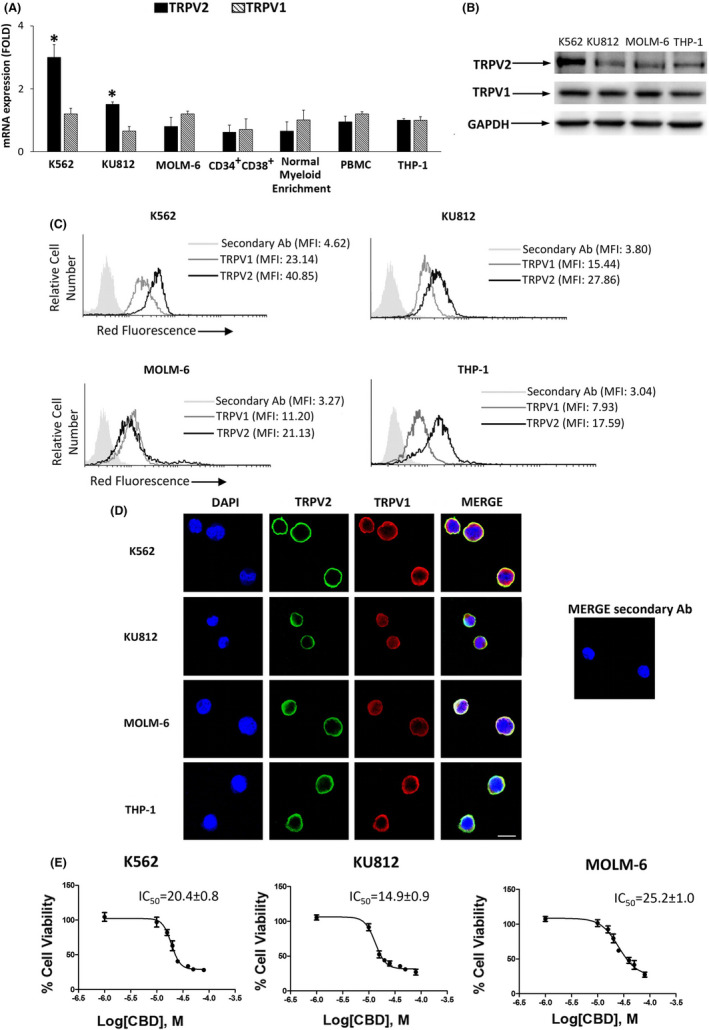
TRPV1 and TRPV2 expression. A, TRPV2 and TRPV1 expression evaluated by qRT‐PCR. Folds (mean ± SD of three separate experiments) = changes respect to THP‐1 cells. *P < .05 vs THP‐1. B, Representative western blot analysis by using anti‐TRPV1 and TRPV2 Abs. GAPDH was used as loading control. C, Cytofluorimetry in CML and THP‐1 cells using anti‐TRPV1 and TRPV2 Abs. MFI, mean fluorescence intensity. D, Confocal microscopy in CML and THP‐1 cells. Bar = 20 µm. Images are representative of three experiments. E, Cell viability in CML cells treated for 24 h with CBD. Data are the mean ± SD of three experiments
Cell viability assay was also performed in CML cells pretreated with 10 μM capsazepine (TRPV1‐antagonist) 22 and 10 μM tranilast (TRPV2‐antagonist) 21 before the addition of CBD for 24 hours. Only tranilast was able to revert the CBD effects (Figure 2A), suggesting the TRPV2 involvement. This was confirmed by using TRPV1 and TRPV2 gene silencing (Figure S1A‐D) and cell viability assay (Figure 2B,C). The CBD‐induced effects were reduced in siTRPV2, but not in siTRPV1, with respect to siGLO cells. Previous findings showed that CBD does not target normal hematopoietic progenitor cells. 23 However, we performed Annexin V/PI staining in PBMCs treated or not with CBD at 25 µM. Data showed that CBD does not induce cell death (Figure S2A,B). Moreover, given that the conventional treatments provoke erythrocyte toxicity, 24 apoptosis and oxidative stress were also evaluated in healthy erythrocytes. No enhancement in the Annexin V fluorescence or ROS production was found in CBD‐respect to vehicle‐treated erythrocytes (Figure S2C‐F).
FIGURE 2.
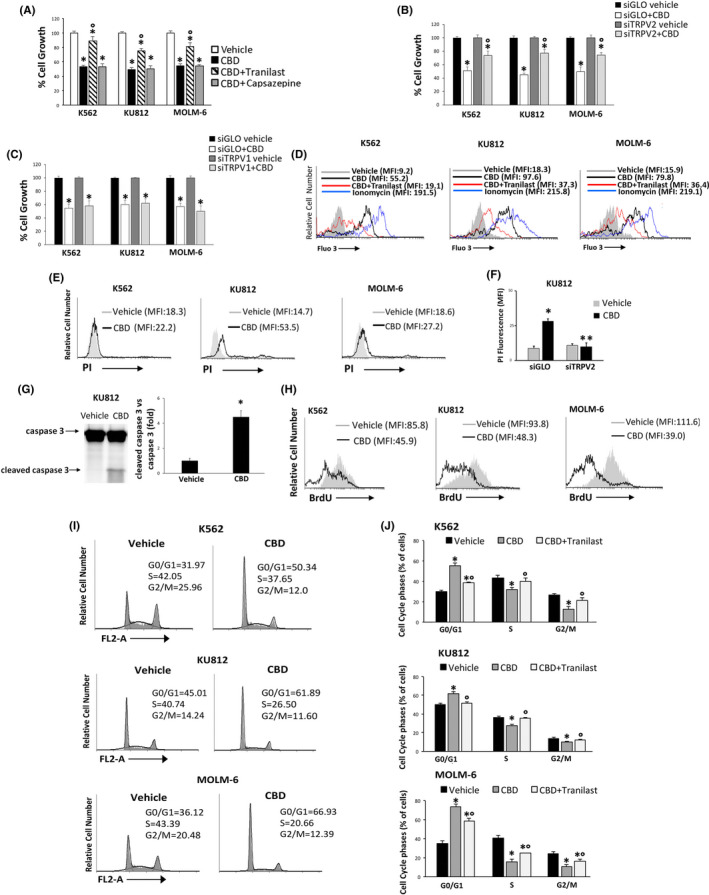
CBD reduces cell viability in a TRPV2‐dependent manner. A, Cell viability in CML cells pretreated with tranilast or capsazepine for 1 h before the addition of CBD at IC50 dose. Vehicle‐treated cells = control. Data are the mean ± SD of three experiments. *P < .05 vs vehicle; °P < .05 vs CBD‐treated cells. B, C, Cell viability in siTRPV2 (B), siTRPV1 (C) and siGLO cells treated for 24 h with CBD at IC50 dose or vehicle. Data are the mean ± SD of three experiments. *P < .05 vs vehicle, °P < .05 vs CBD‐treated siGLO cells. D, [Ca2+]i evaluated by Fluo‐3 staining and FACS analysis in CML cells treated for 1 min with CBD (IC50 dose) or pretreated with tranilast and then stimulated with CBD. E, PI staining in CML cells treated with CBD at IC50 dose or with vehicle for 24 h. Data are representative of three experiments. F, PI assay performed in CBD (IC50 dose)‐ or vehicle‐treated for 24 h siGLO and siTRPV2 KU812 cells. Data are the mean ± SD of three experiments. *P < .05 vs vehicle; **P < .05 vs CBD‐treated siGLO. G, Caspase 3 analyzed by western blot in KU812 cells treated as described in panel E. Blot is representative of three experiments. For statistical analysis, cleaved caspase 3 densitometry values were normalized to caspase 3 levels. Folds represent changes respect to vehicle. H, BrdU assay in CML cells treated with CBD at IC50 dose or with vehicle for 24 h. Histograms are representative of three experiments. I, Cell cycle analysis in CML cells treated with CBD at IC50 dose or with vehicle for 24 h. Numbers = %cells in each cell cycle step. Data are representative of three experiments. J, Statistical analysis of cell cycle phases in CML cells treated with CBD at IC50 dose or vehicle for 24 h or pretreated with tranilast for 1 h before the addition of CBD. Data are the mean ± SD of three experiments. *P <.05 vs vehicle, °P <.05 vs CBD
Since TRPV2 is a calcium permeable channel, 9 we performed [Ca2+]i assay up to 3 minutes (data not shown). Our results showed that CBD induced a rapid increase in [Ca2+]i 1 minute after the stimulation. This effect was inhibited by tranilast supporting the TRPV2 involvement (Figures 2D and S2G). Moreover, the calcium overload was associated with a marked enhancement in ROS production after 1 hour of CBD treatment (Figure S2H,I).
3.2. CBD inhibits cell proliferation
Cytofluorimetric analysis, performed in CML cells treated for 24 hours with CBD and stained with PI, demonstrated that CBD mildly stimulates cell death only in KU812 cells. CDB‐induced cell death was reverted in siTRPV2 KU812 cells, underlying the involvement of this channel (Figures 2E,F and S3A). The presence of cleaved caspase 3 fragment in CBD‐treated KU812 cells was found indicating apoptosis (Figure 2G). Moreover, CBD markedly inhibited BrdU incorporation (Figures 2H, and S3B) in all CML lines indicating that the reduction in cell viability is associated with a strong decrease in cell proliferation. CBD treatment also induced a TRPV2‐dependent cell cycle arrest by increasing the percentage of cells in the G0/G1 phase (Figure 2I,J).
3.3. The CBD‐induced effects are associated with mitochondria impairment
Calcium overload and ROS production play a pivotal role in the mitochondria dysfunction. 25 Thus, by using JC‐1 staining, we found that the treatment of CML cells with the IC50 dose of CBD induces a time‐dependent ΔΨm that was evident at 8 hours, increased at 12 hours, and slightly decreased at 24 hours (Figures 3A and S4A). In PBMCs treated with the highest dose of CBD, data showed the absence of cells with depolarized mitochondria (decrease in red fluorescence) (Figure S4B). To further support the role of TRPV2, we evaluated the drop of ΔΨm in siGLO and siTRPV2 cells treated with vehicle or with CBD (IC50 dose) for 12 hours. Our results showed that the CBD‐induced mitochondrial impairment is markedly reduced in siTRPV2 respect to siGLO cells (Figure 3B). Moreover, a marked reduction in Mitobright fluorescence and an increased cell percentage with diminished green fluorescence in CBD‐ respect to vehicle‐treated cells were found, indicating mitochondrial impairment (Figures 3C and S4C).
FIGURE 3.
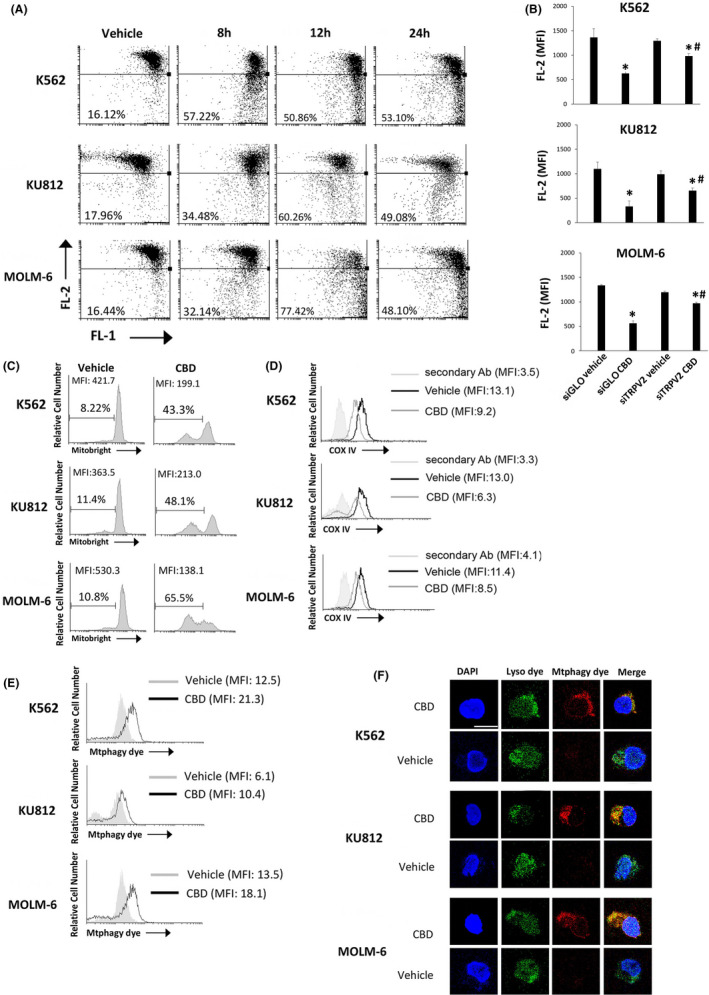
CBD impairs mitochondria. A, ΔΨm changes on CBD‐treated CML cells by JC‐1 staining and FACS. Drop of ∆Ψm decreases the J‐aggregate (red fluorescence). Numbers = %cells showing drop of ΔΨm. Data are representative of three experiments. B, Drop of ∆Ψm evaluated by JC‐1 in siGLO and siTRPV2 cells treated with vehicle or CBD for 12 h. Data are the mean ± SD of three experiments *P < .05 vs vehicle; # P < .05 vs CBD‐treated siGLO cells. C, Cytofluorimetric analysis of CML cells treated with CBD at IC50 dose or with vehicle for 24 h and labeled with Mitobright. Histograms are representative of three experiments. D, COX IV expression in CML cells, treated for 24 h as above described, investigated by cytofluorimetry. Data are representative of three experiments. E, Mtphagy dye analysis in CML cells after 24 h of CBD treatment. Data are representative of three experiments. F, Confocal microscopy of CML cells treated as above described and stained with Mtphagy/Lyso dyes. Images are representative of three experiments. Bar: 20 µm
3.4. CBD induces mitophagy in CML cells
We then labeled cells with the specific anti‐COX IV Ab, used as mitochondrial mass marker. 26 A marked down‐regulation in COX IV expression in CBD‐ respect to vehicle‐treated CML cells was found demonstrating the mitochondria removal (Figures 3D and S4D). Thus, mitophagy assay was performed. An increase of the Mtphagy dye fluorescence in CBD‐ respect to vehicle‐treated CML cells was found as shown by the MFI values (Figures 3E and S4E). To corroborate the fusion of Mtphagy dye‐labeled mitochondria with lysosomes, cells were also stained with Lyso dye and then analyzed by confocal microscopy. The analysis confirmed the strongly enhancement in Mtphagy dye fluorescence and the colocalization between Mtphagy and Lyso dyes in CBD‐ respect to vehicle‐treated cells indicating that damaged mitochondria are driven inside lysosomes (Figure 3F).
3.5. The CBD‐induced mitophagy is TRPV2‐dependent in CML cells
Mitophagy is a cargo‐specific autophagy 27 so we examined the conversion of the soluble form of LC3‐I to the lipidated and autophagosome‐associated form (LC3‐II).
CBD treatment increased the expression of LC3‐II in all CML cell lines (Figure 4A). The autophagy inhibitor BAF, by blocking the autophagic degradation activity, increased the LC3‐II form (Figure S4F) demonstrating that LC3‐II levels were dependent from the autophagic flux and not the result of protein synthesis. In addition, in siTRPV2 cells, CBD was unable to markedly increase the expression of LC3‐II respect to vehicle‐treated cells with comparable level detected in siGLO cells, supporting the involvement of TRPV2 (Figure 4B). CBD induced up‐regulation of ATG16L1 and ATG5/ATG12 expression indicating the execution of autophagy (Figure 4C). 28 Given that mitophagy of optineurin‐labeled mitochondria is regulated by parkin and PINK1, 29 we evaluated the expression of these proteins in CML cells treated or not with CBD. Our findings showed that CBD stimulates an increased expression of parkin, PINK1, and optineurin (Figure 4D).
FIGURE 4.
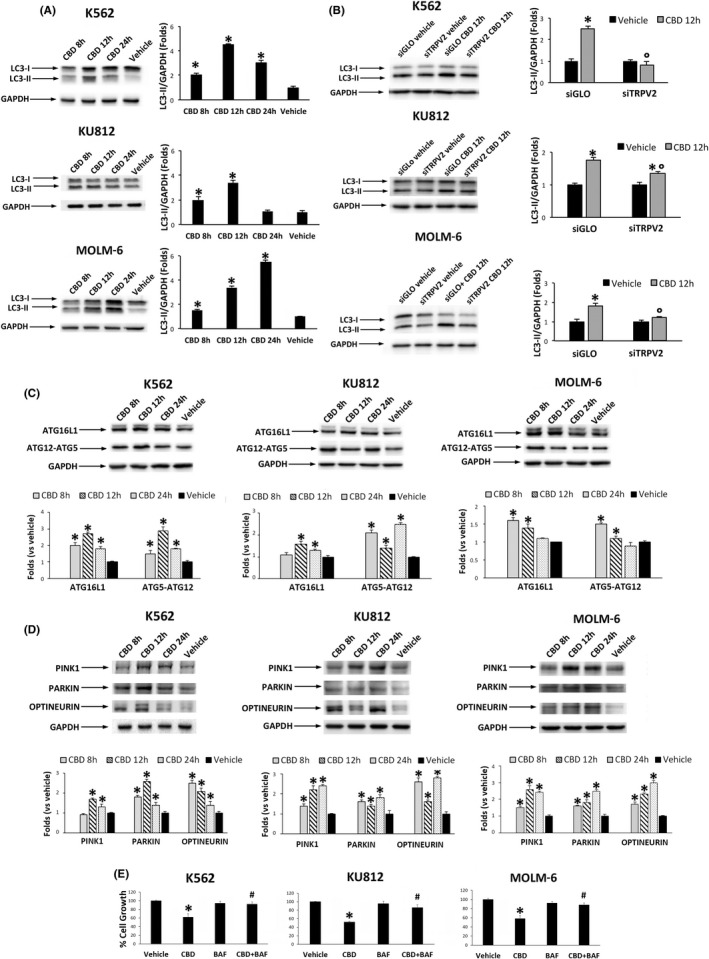
CBD increases the expression of mitophagy markers. A, Western blot analysis of LC‐3 II in CML cells treated for different times with CBD at IC50 dose or with vehicle. *P < .01 vs vehicle. B, Western blot analysis of LC‐3 II in siGLO and siTRPV2 CML cells treated with CBD at IC50 dose or with vehicle for 12 h. *P < .01 vs vehicle; °P < .05 vs CBD‐treated siGLO cells. C, Western blot analysis of ATG16L‐1 and ATG12‐ATG5 complex in CML cells treated with CBD at IC50 dose for different times. *P < .01 vs vehicle. D, Western blot analysis of pink1, parkin, and optineurin in CML cells treated with CBD at IC50 dose for different times. *P < .01 vs vehicle. Blots are representative of three experiments. GAPDH was used as loading control. Folds (mean ± SD of three experiments) = changes respect to vehicle. E, Cell viability assay performed in CML cells pretreated for 1 h with BAF and then treated for 24 h with CBD (IC50 dose). *P < .05 vs vehicle‐treated cells; **P < .05 vs CBD‐treated cells
Moreover, the inhibition of autophagy strongly reverted the CBD‐induced cell growth inhibition (Figure 4E) supporting the role of autophagy in the CBD‐mediated effects.
Autophagy can modulate the levels of pluripotency‐associated proteins. 30 Thus, we investigated the expression of transcription factors involved in the maturation of CML cells. CBD reduced the expression of the stemness‐related protein OCT‐4, comparing CBD‐ respect to vehicle‐treated cells (Figure 5A,B). By western blot analysis and confocal microscopy, we also demonstrated that CBD increases the expression of PU.1, essential for myeloid cell development 31 indicating an enhancement in CML cell maturation (Figure 5C,D). The ability of CBD to stimulate a more differentiated state was also evaluated by colony forming assay, useful to test the clonogenic activity of undifferentiated cells. 32 We demonstrated that CBD strongly reduces the ability to form colonies in all CML cell lines (Figure 5E,F).
FIGURE 5.
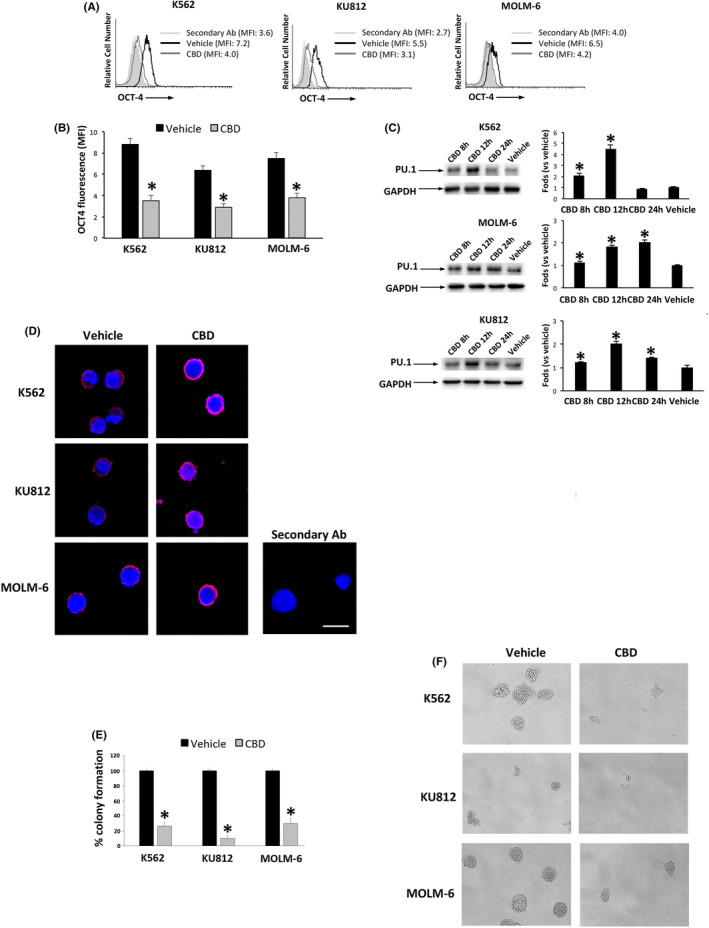
CBD changes the expression of differentiation markers. A, OCT‐4 expression by FACS in CML cells treated for 24 h with CBD at IC50 dose or vehicle. Data are representative of three separate experiments. MFI, mean fluorescence intensity. B, Statistical analysis of the MFI values of panel A. Data are the mean ± SD of three experiments. *P < .05 vs vehicle. C, Western blot analysis of PU.1 in CML cells treated with CBD at IC50 dose or vehicle for different times. Blots are representative of three experiments. GAPDH was used as loading control. Folds represent changes respect to vehicle. Data are the mean ± SD of three experiments. *P < .05 vs vehicle. D, PU.1 expression by confocal microscopy. Images are representative of three experiments. Bar = 20 µm. E, Colony formation assay in CML cells treated with CBD (IC50 dose) for 24 h and then plated in Methocult for 10 d. Data are the mean ± SD of three experiments. *P < .05 vs vehicle‐treated cells. F, Representative images of the colony formation in CML cells treated as above described. Magnification = 10×
3.6. CBD synergizes with imatinib in reducing cell viability in CML cell lines
Currently, the standard therapy of CML consists of TKIs such as imatinib. We evaluated the possible synergism between CBD and imatinib in all CML cell lines. Cells were exposed to various concentrations of CBD and imatinib for 24 hours. The isobologram analysis showed that several combinations of the two drugs induce increased levels of cytotoxicity, as compared with single treatments (Figure 6A).
FIGURE 6.
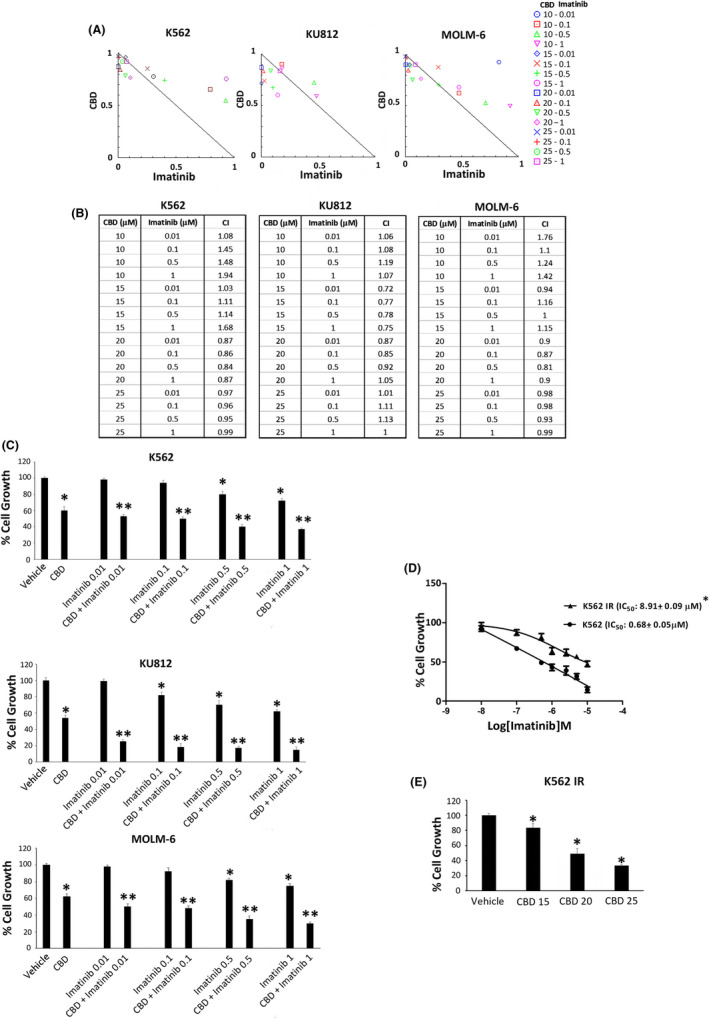
CBD and imatinib synergize in reducing cell viability. A, Isobologram plots for combination treatments of CBD and imatinib. Lower left of the hypotenuse, synergism; hypotenuse, additive effect; upper right of the hypotenuse, antagonism. Data are representative of three experiments. B, CI values by CompuSyn software. Data are representative of three experiments. C, Cell viability assay in CML cells treated with CBD at IC50 dose and imatinib at different doses for 24 h. Data are the mean ± SD of three experiments. *P < .01 vs vehicle; **P < .01 vs CBD or imatinib. D, Cell viability assay performed in K562 IR or K562 cells treated with imatinib or vehicle. *P < .01 vs K562 cells. E, Cell viability assay in K562 IR treated with CBD at IC50 doses for 24 h. Data are the mean ± SD of three experiments. *P < .01 vs vehicle
The CI values obtained by combining the CBD at IC50 dose with imatinib are <1, indicating from slight to moderate synergistic effects (Figure 6B). In fact, the percentage of cell growth was markedly reduced when imatinib, even at the lowest dose, is combined with CBD at IC50 dose (Figure 6C). To strengthen these data, we also performed cell viability assay in K562 IR treated with different doses of CBD for 24 hours. After confirmation of the resistant phenotype in K562 IR (Figure 6D), as shown in Figure 6E, we found that CBD treatment also inhibits cell growth in this resistant cell model.
4. DISCUSSION
Chronic myeloid leukemia is a myeloproliferative disorder originating from an incomplete differentiation process of hematopoietic progenitor to adult cells with consequent accumulation of immature ones. Although successful results were obtained with TKIs in therapy, several patients show or develop resistance. 1 , 2 Thus, the discovery of new anticancer compounds and the identification of new drug targets represent an interesting challenge.
CBD is considered a promising anticancer drug alone or in combinations with conventional chemotherapies. In fact, findings demonstrated that CBD induces cell death and inhibits cell migration and vascularization in glioma models, and similar results have been obtained in other cancer types. 33 , 34 , 35
Few data on CBD‐mediated effects in leukemia and lymphoma have been provided 1 , 2 , 36 , 37 , 38 and no findings are present about the role of TRPV2 in CBD‐induced effects on CML cells.
Although CBD belongs to the family of phytocannabinoids able to bind the CB1 and CB2 receptors, the idea that CBD has no affinity for these receptors is gaining strength. 11 Instead, it has been accepted that it is an agonist for TRPV1 and TRPV2. 20 , 21 TRPV1 and TRPV2, ion channels involved in the regulation of calcium signaling, are expressed on normal immune cells 39 , 40 and in some leukemia cells. 9 , 19 , 41 , 42 Our data support the expression of both receptors in the three CML cell lines in common myeloid progenitors and normal myeloid cells, suggesting the importance of TRPV channels in the modulation of immune activities. In particular, TRPV2 is implicated phagocytosis, degranulation, chemotaxis, cytokine secretion, and proliferation. Little is known about the TRPV2 role in the leukemogenesis. An oncogenic function related to its involvement in cell proliferation has been suggested. 9
Although TRPV2 expression is higher in K562 respect to KU812 and MOLM‐6 cells, the ability of CBD to inhibit cell growth is very similar among the three CML lines. mRNA and protein expression levels are not enough to suppose differences in the TRPV functionality. In fact, the status of TRPV2 activation is influenced by post‐translational modifications such as glycosylation, phosphorylation, and binding with lipids. 43
Interestingly, we demonstrated that CBD, by activating TRPV2, induces inhibition of cell proliferation and blockage of the cell cycle. Also, our previous conclusions in glioma cells indicating that the CBD‐induced effects are TRPV2‐dependent 21 suggest its relevant role in the CBD‐stimulated signaling pathways.
We showed for the first time that the CBD‐induced TRPV2 dependent inhibition of cell proliferation is associated with mitophagy in CML cells. The ability of CBD to cause mitochondrial damage and to interact directly with isolated mitochondria was recently investigated in Jurkat cells. 12 Here we demonstrated that CBD stimulates, via TRPV2, calcium overload, and oxidative stress strongly leading to the mitochondrial dysfunction. 25 Mitochondria dynamics is essential to regulate cellular metabolism, ATP production, and redox homeostasis. 13 It has been found that the number of mitochondria and the total mitochondrial mass are increased in chronic lymphatic leukemia with respect to normal naïve B lymphocytes. 44 Moreover, the targeting of mitochondrial enzymes represents an effective therapeutic strategy for leukemia cells since they have more mitochondria compared with normal hematopoietic stem cells. 45 The upregulation of mitochondrial mass associated with chemoresistance has been found in lymphoma. 46 Thus, our data showing that CBD treatment induces mitochondria dysfunction and promotes mitophagy meet the overall need to use therapies inhibiting exaggerated mitochondrial activity in leukemia. 13
Mitophagy is responsible for the removal of damaged mitochondria. 27 , 38 ATG family proteins such as LC3, ATG5, ATG12, and ATG16‐L1 regulate the autophagy by participating in autophagosome formation. 47 , 48 In our study, the CBD treatment, via TRPV2, increased the expression of LC3‐II. It also enhanced ATG16‐L1 and ATG5‐ATG12 complex levels, indicating the stimulation of the autophagy. These results are in agreement with previous findings demonstrating the upregulation of these proteins after inducing photodamages in mitochondria. The pink1 and parkin upregulation and accumulation on mitochondria are essential to promote mitophagy. 49 Mitophagy is also regulated by optineurin that binds damaged mitochondria and increases their autophagic engulfment. 31 Similarly, we demonstrated that CBD enhances the expression of pink1, parkin, and optineurin.
Although findings suggest that mitophagy plays a role in the maintenance of cancer stem cells, 50 it is also well accepted that mitophagy participates in cellular differentiation. 51 In agreement, we showed that cell cycle arrest and mitophagy induced by CBD in CML cells are associated with changes in the differentiation markers OCT‐4 and PU.1 involved in leukemogenesis and myeloid differentiation. 52 , 53 We found a reduction of the stem‐associated marker OCT‐4 level and the enhancement of PU.1, suggesting that CBD drives CML cells to a more differentiated state. This was also confirmed by colony formation assay that demonstrates the ability of CBD treatment to reduce the clonogenic activity of CML cells.
The presence of poorly differentiated cells is associated with chemoresistance. TKI resistance develops due to secondary BCR‐ABL mutations and/or an independent mechanism. 54 Several studies have suggested that TKI treatment, surprisingly, induces stemness in CML cells, 55 thus the need to develop new targeted therapies that could overcome these stemness‐promoting effects is attractive. In this regard, we highlighted the ability of CBD to act synergistically with imatinib and reduce the cell viability of IR cells. Overall, given that CBD is characterized by a favorable safety and tolerability profile, 56 the use of CBD as adjuvant to enhance conventional therapy and improve the prognosis of patients by modulating TRPV2 activity is very promising in CML clinical therapy.
DISCLOSURE
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supporting information
Fig S1
{kind=link}
Fig S2
{kind=link}
Fig S3
{kind=link}
Fig S4
{kind=link}
Figure Captions
Maggi F, Morelli MB, Tomassoni D, et al. The effects of cannabidiol via TRPV2 channel in chronic myeloid leukemia cells and its combination with imatinib. Cancer Sci. 2022;113:1235–1249. doi: 10.1111/cas.15257
Federica Maggi and Maria Beatrice Morelli contributed equally to this work.
Funding information
PRIN 2017 (prot. 2017FS5SHL), Italian Ministry of Education, University and Research; Progetti per Avvio alla Ricerca (n. protocollo AR120172B93FBB5B) Sapienza University; Progetto FAR 2018, University of Camerino
DATA AVAILABILITY STATEMENT
Data supporting the conclusions of this article will be made available by the authors.
REFERENCES
- 1. Osman AEG, Deininger MW. Chronic myeloid leukemia: modern therapies, current challenges and future directions. Blood Rev. 2021;49:100825. doi: 10.1016/j.blre.2021.100825 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Apperley JF. Chronic myeloid leukaemia. Lancet. 2015;385:1447‐1459. doi: 10.1016/S0140-6736(13)62120-0 [DOI] [PubMed] [Google Scholar]
- 3. Shapovalov G, Ritaine A, Skryma R, Prevarskaya N. Role of TRP ion channels in cancer and tumorigenesis. Semin Immunopathol. 2016;38:357‐369. doi: 10.1007/s00281-015-0525-1 [DOI] [PubMed] [Google Scholar]
- 4. Zhao Y, McVeigh BM, Moiseenkova‐Bell VY. Structural pharmacology of TRP channels. J Mol Biol. 2021;433(17):166914. doi: 10.1016/j.jmb.2021.166914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Morelli MB, Liberati S, Amantini C, et al. Expression and function of the transient receptor potential ion channel family in the hematologic malignancies. Curr Mol Pharmacol. 2013;6:137‐148. [DOI] [PubMed] [Google Scholar]
- 6. Maggi F, Morelli MB, Nabissi M, et al. Transient receptor potential (TRP) channels in haematological malignancies: an update. Biomolecules. 2021;11:765. doi: 10.3390/biom11050765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Heise N, Palme D, Misovic M. Non‐selective cation channel‐mediated Ca2+‐entry and activation of Ca2+/calmodulin‐dependent kinase II contribute to G2/M cell cycle arrest and survival of irradiated leukemia cells. Cell Physiol Biochem. 2010;26:597‐608. [DOI] [PubMed] [Google Scholar]
- 8. Takahashi K, Umebayashi C, Numata T, et al. TRPM7‐mediated spontaneous Ca2+ entry regulates the proliferation and differentiation of human leukemia cell line K562. Physiol Rep. 2018;6:1‐15. doi: 10.14814/phy2.13796 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Siveen KS, Prabhu KS, Parray AS, et al. Evaluation of cationic channel TRPV2 as a novel biomarker and therapeutic target in leukemia‐implications concerning the resolution of pulmonary inflammation. Sci Rep. 2019;9:1554. doi: 10.1038/s41598-018-37469-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Meotti FC, Lemos de Andrade E, Calixto JB. TRP modulation by natural compounds. Handb Exp Pharmacol. 2014;223:1177‐1238. doi: 10.1007/978-3-319-05161-1_19 [DOI] [PubMed] [Google Scholar]
- 11. Stasiłowicz A, Tomala A, Podolak I, Cielecka‐Piontek J Cannabis sativa L. as a natural drug meeting the criteria of a multitarget approach to treatment. Int J Mol Sci. 2021;22:778. doi: 10.3390/ijms22020778 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Olivas‐Aguirre M, Torres‐López L, Valle‐Reyes JS, Hernández‐Cruz A, Pottosin I, Dobrovinskaya O. Cannabidiol directly targets mitochondria and disturbs calcium homeostasis in acute lymphoblastic leukemia. Cell Death Dis. 2019;10:779. doi: 10.1038/s41419-019-2024-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Barbato A, Scandura G, Puglisi F, et al. Mitochondrial bioenergetics at the onset of drug resistance in hematological malignancies: an overview. Front Oncol. 2020;10:604143. doi: 10.3389/fonc.2020.604143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Dong Y, Gao X, Zhao Y, et al. Semi‐random mutagenesis profile of BCR‐ABL during imatinib resistance acquirement in K562 cells. Mol Med Rep. 2017;16:9409‐9414. doi: 10.3892/mmr.2017.7835 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Dircio‐Maldonado R, Flores‐Guzman P, Corral‐Navarro J, et al. Functional integrity and gene expression profiles of human cord blood‐derived hematopoietic stem and progenitor cells generated in vitro. Stem Cells Transl Med. 2018;7:602‐614. doi: 10.1002/sctm.18-0013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. de Abreu Costa L, Henrique Fernandes Ottoni M, dos Santos M, et al. Dimethyl sulfoxide (DMSO) decreases cell proliferation and TNF‐α, IFN‐γ, and IL‐2 cytokines production in cultures of peripheral blood lymphocytes. Molecules. 2017;22:1789. doi: 10.3390/molecules22111789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Khanduja KL, Avti PK, Kumar S, Mittal N, Sohi KK, Pathak CM. Anti‐apoptotic activity of caffeic acid, ellagic acid and ferulic acid in normal human peripheral blood mononuclear cells: a Bcl‐2 independent mechanism. Biochim Biophys Acta. 2006;1760:283‐289. doi: 10.1016/j.bbagen.2005.12.017 [DOI] [PubMed] [Google Scholar]
- 18. Sun Y, Liu G, Jiang Y, Wang H, Xiao H, Guan G. Erythropoietin protects erythrocytes against oxidative stress‐induced eryptosis in vitro. Clin Lab. 2018;64: doi: 10.7754/Clin.Lab.2017.170924 [DOI] [PubMed] [Google Scholar]
- 19. Kunde DA, Yingchoncharoen J, Jurković S, Geraghty DP. TRPV1 mediates capsaicin‐stimulated metabolic activity but not cell death or inhibition of interleukin‐1β release in human THP‐1 monocytes. Toxicol Appl Pharmacol. 2018;360:9‐17. doi: 10.1016/J.TAAP.2018.09.025 [DOI] [PubMed] [Google Scholar]
- 20. Iannotti FA, Hill CL, Leo A, et al. Nonpsychotropic plant cannabinoids, cannabidivarin (CBDV) and cannabidiol (CBD), activate and desensitize transient receptor potential vanilloid 1 (TRPV1) channels in vitro: potential for the treatment of neuronal hyperexcitability. ACS Chem Neurosci. 2014;5:1131‐1141. doi: 10.1021/cn5000524 [DOI] [PubMed] [Google Scholar]
- 21. Nabissi M, Morelli MB, Amantini C, et al. Cannabidiol stimulates Aml‐1a‐dependent glial differentiation and inhibits glioma stem‐like cells proliferation by inducing autophagy in a TRPV2‐dependent manner. Int J Cancer. 2015;137:1855‐1869. doi: 10.1002/ijc.29573 [DOI] [PubMed] [Google Scholar]
- 22. Amantini C, Mosca M, Lucciarini R, et al. Distinct thymocyte subsets express the vanilloid receptor VR1 that mediates capsaicin‐induced apoptotic cell death. Cell Death Differ. 2004;11:1342‐1356. doi: 10.1038/sj.cdd.4401506 [DOI] [PubMed] [Google Scholar]
- 23. Morelli MB, Offidani M, Alesiani F, et al. The effects of cannabidiol and its synergism with bortezomib in multiple myeloma cell lines. A role for transient receptor potential vanilloid type‐2. Int J Cancer. 2014;134:2534‐2546. doi: 10.1002/ijc.28591 [DOI] [PubMed] [Google Scholar]
- 24. Barber NA, Afzal W, Akhtari M. Hematologic toxicities of small molecule tyrosine kinase inhibitors. Target Oncol. 2011;6:203‐215. doi: 10.1007/s11523-011-0202-9 [DOI] [PubMed] [Google Scholar]
- 25. Brookes PS, Yoon Y, Robotham JL, Anders MW, Sheu S‐S. Calcium, ATP, and ROS: a mitochondrial love‐hate triangle. Am J Physiol Cell Physiol. 2004;287:C817‐C833. doi: 10.1152/ajpcell.00139.2004 [DOI] [PubMed] [Google Scholar]
- 26. Dannheisig DP, Beck E, Calzia E, Walther P, Behrends C, Pfister AS. Loss of Peter Pan (PPAN) affects mitochondrial homeostasis and autophagic flux. Cells. 2019;8:894. doi: 10.3390/cells8080894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Youle RJ, Narendra DP. Mechanisms of mitophagy. Nat Rev Mol Cell Biol. 2011;12:9‐14. doi: 10.1038/nrm3028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Kharaziha P, Panaretakis T. Dynamics of Atg5–Atg12–Atg16L1 aggregation and deaggregation. Methods Enzymol. 2017;587:247‐255. doi: 10.1016/bs.mie.2016.09.059 [DOI] [PubMed] [Google Scholar]
- 29. Wong YC, Holzbaur ELF. Optineurin is an autophagy receptor for damaged mitochondria in parkin‐mediated mitophagy that is disrupted by an ALS‐linked mutation. Proc Natl Acad Sci USA. 2014;111:E4439‐E4448. doi: 10.1073/pnas.1405752111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Cho Y‐H, Han K‐M, Kim D, et al. Autophagy regulates homeostasis of pluripotency‐associated proteins in hESCs. Stem Cells. 2014;32:424‐435. doi: 10.1002/stem.1589 [DOI] [PubMed] [Google Scholar]
- 31. Albajar M, Gutierrez P, Richard C, et al. PU.1 expression is restored upon treatment of chronic myeloid leukemia patients. Cancer Lett. 2008;270:328‐336. doi: 10.1016/j.canlet.2008.05.024 [DOI] [PubMed] [Google Scholar]
- 32. Rajendran V, Jain MV. In vitro tumorigenic assay: colony forming assay for cancer stem cells. Methods Mol Biol. 2018;1692:89‐95. doi: 10.1007/978-1-4939-7401-6_8 [DOI] [PubMed] [Google Scholar]
- 33. Seltzer ES, Watters AK, MacKenzie D, Granat LM, Zhang D. Cannabidiol (CBD) as a promising anti‐cancer drug. Cancers. 2020;12:3203. doi: 10.3390/cancers12113203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Solinas M, Massi P, Cinquina V, et al. Cannabidiol, a non‐psychoactive cannabinoid compound, inhibits proliferation and invasion in U87‐MG and T98G glioma cells through a multitarget effect. PLoS One. 2013;8:e76918. doi: 10.1371/journal.pone.0076918 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Deng L, Ng L, Ozawa T, Stella N. Quantitative analyses of synergistic responses between cannabidiol and DNA‐damaging agents on the proliferation and viability of glioblastoma and neural progenitor cells in culture. J Pharmacol Exp Ther. 2017;360:215‐224. doi: 10.1124/jpet.116.236968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Kalenderoglou N, Macpherson T, Wright KL. Cannabidiol reduces leukemic cell size – but is it important? Front Pharmacol. 2017;8:144. doi: 10.3389/fphar.2017.00144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Scott KA, Dalgleish AG, Liu WM. Anticancer effects of phytocannabinoids used with chemotherapy in leukaemia cells can be improved by altering the sequence of their administration. Int J Oncol. 2017;51:369‐377. doi: 10.3892/ijo.2017.4022 [DOI] [PubMed] [Google Scholar]
- 38. Togano T, Kim N, Kim N, et al. The evaluation of cannabidiol's effect on the immunotherapy of Burkitt lymphoma. Biochem Biophys Res Comm. 2019;520:225‐230. doi: 10.1016/j.bbrc.2019.10.001 [DOI] [PubMed] [Google Scholar]
- 39. Amantini C, Farfariello V, Cardinali C, et al. The TRPV1 ion channel regulates thymocyte differentiation by modulating autophagy and proteasome activity. Oncotarget. 2017;8:90766‐90780. doi: 10.18632/oncotarget.21798 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Cai X, Yang Y‐C, Wang J‐F, et al. Transient receptor potential vanilloid 2 (TRPV2), a potential novel biomarker in childhood asthma. J Asthma. 2013;50:209‐214. doi: 10.3109/02770903.2012.753454 [DOI] [PubMed] [Google Scholar]
- 41. Punzo F, Manzo I, Tortora C, et al. Effects of CB2 and TRPV1 receptors' stimulation in pediatric acute T‐lymphoblastic leukemia. Oncotarget. 2018;9:21244‐21258. doi: 10.18632/oncotarget.25052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Pottosin I, Delgado‐Enciso I, Bonales‐Alatorre E, Nieto‐Pescador MG, Moreno‐Galindo EG, Dobrovinskaya O. Mechanosensitive Ca2 +‐permeable channels in human leukemic cells: pharmacological and molecular evidence for TRPV2. Biochim Biophys Acta. 2015;1848:51‐59. doi: 10.1016/j.bbamem.2014.09.008 [DOI] [PubMed] [Google Scholar]
- 43. Huynh KW, Cohen MR, Jiang J, et al. Structure of the full‐length TRPV2 channel by cryo‐EM. Nat Commun. 2016;7:11130. doi: 10.1038/ncomms11130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Nakajima K, Kawashima I, Koshiisi M, et al. Glycolytic enzyme hexokinase II is a putative therapeutic target in B‐cell malignant lymphoma. Exp Hematol. 2019;78:46‐55.e3. doi: 10.1016/j.exphem.2019.09.023 [DOI] [PubMed] [Google Scholar]
- 45. Sriskanthadevan S, Jeyaraju DV, Chung TE, et al. AML cells have low spare reserve capacity in their respiratory chain that renders them susceptible to oxidative metabolic stress. Blood. 2015;125:2120‐2130. doi: 10.1182/blood-2014-08-594408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Mikkilineni L, Whitaker‐Menezes D, Domingo‐Vidal M, et al. Hodgkin lymphoma: a complex metabolic ecosystem with glycolytic reprogramming of the tumor microenvironment. Semin Oncol. 2017;44:218‐225. doi: 10.1053/j.seminoncol.2017.10.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Tanida I, Ueno T, Kominami E. LC3 conjugation system in mammalian autophagy. Int J Biochem Cell Biol. 2004;36:2503‐2518. doi: 10.1016/j.biocel.2004.05.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Hanada T, Noda NN, Satomi Y, et al. The Atg12‐Atg5 conjugate has a novel E3‐like activity for protein lipidation in autophagy. J Biol Chem. 2007;282:37298‐37302. doi: 10.1074/jbc.C700195200 [DOI] [PubMed] [Google Scholar]
- 49. Narendra DP, Youle RJ. Targeting mitochondrial dysfunction: role for PINK1 and Parkin in mitochondrial quality control. Antioxid Redox Signal. 2011;14:1929‐1938. doi: 10.1089/ars.2010.3799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Lévêque M, Penna A, Le Trionnaire S, et al. Phagocytosis depends on TRPV2‐mediated calcium influx and requires TRPV2 in lipids rafts: alteration in macrophages from patients with cystic fibrosis. Sci Rep. 2018;8:4310. doi: 10.1038/s41598-018-22558-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Naik PP, Birbrair A, Bhutia SK. Mitophagy‐driven metabolic switch reprograms stem cell fate. Cell Mol Life Sci. 2019;76:27‐43. doi: 10.1007/s00018-018-2922-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Picot T, Kesr S, Wu Y, et al. Potential role of OCT4 in leukemogenesis. Stem Cells Dev. 2017;26:1637‐1647. doi: 10.1089/scd.2017.0134 [DOI] [PubMed] [Google Scholar]
- 53. Trinh BQ, Ummarino S, Zhang Y, et al. Myeloid lncRNA LOUP mediates opposing regulatory effects of RUNX1 and RUNX1‐ETO in t(8;21) AML. Blood. 2021;138:1331‐1344. doi: 10.1182/blood.2020007920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Arrigoni E, Del Re M, Galimberti S, et al. Concise review: chronic myeloid leukemia: stem cell niche and response to pharmacologic treatment. Stem Cells Transl Med. 2018;7:305‐314. doi: 10.1002/sctm.17-0175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Charaf L, Mahon F‐X, Lamrissi‐Garcia I, et al. Effect of tyrosine kinase inhibitors on stemness in normal and chronic myeloid leukemia cells. Leukemia. 2017;31:65‐74. doi: 10.1038/leu.2016.154 [DOI] [PubMed] [Google Scholar]
- 56. Millar SA, Stone NL, Bellman ZD, Yates AS, England TJ, O'Sullivan SE. A systematic review of cannabidiol dosing in clinical populations. Br J Clin Pharmacol. 2019;85:1888‐1900. doi: 10.1111/bcp.14038 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig S1
Fig S2
Fig S3
Fig S4
Figure Captions
Data Availability Statement
Data supporting the conclusions of this article will be made available by the authors.