
Fig. 1.
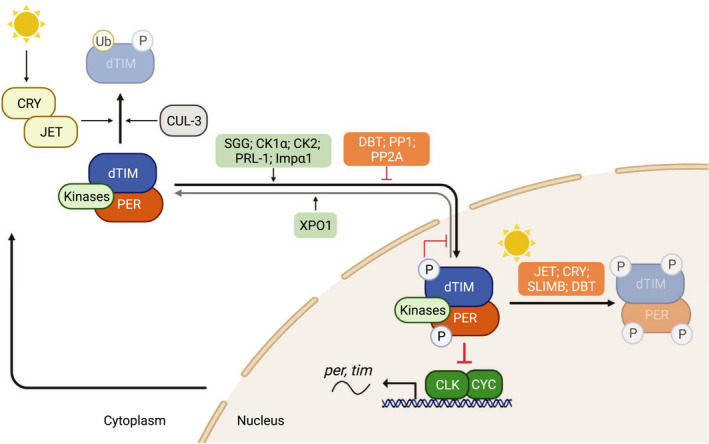
Drosophila TIM (dTIM) is a core component of the molecular oscillator. During the day, CLK‐CYC heterodimers activate the transcription of rhythmic genes, including per and tim in the nucleus [6]. In the cytoplasm, dTIM undergoes proteasomal degradation mediated by CRYPTOCHROME (CRY) [35, 36, 37, 38] and JETLAG (JET) [28, 88] upon light exposure. CULLIN‐3 (CUL‐3) has also been observed to mediate dTIM degradation in a light‐independent manner [27]. Early in the night, SHAGGY (SGG) [68], casein kinase 1α (CK1α) [187], casein kinase 2 (CK2) [69, 70], Importin‐α1 (Impα1) [64] and phosphatase of regenerating liver‐1 (PRL‐1) [75] promote nuclear accumulation of PER‐dTIM complex. This is antagonized by DOUBLETIME (DBT) [25], protein phosphatase 1 (PP1) [74] and protein phosphatase 2A (PP2A) [73]. Once PER‐dTIM complex is in the nucleus, CK2‐dependent phosphorylation of dTIM (S1404) inhibits PER‐dTIM nuclear export by exportin 1 (XPO1) complex, retaining PER‐dTIM complex in the nucleus [67]. At midnight, nuclear PER–dTIM complex interacts with CLK‐CYC and represses their transcriptional activity [23, 25]. From late night to early morning, CRY and JET mediate light‐dependent TIM degradation [28, 88], whereas DBT and SUPERNUMERARY LIMBS (SLIMB) mediate PER degradation [26, 29]. There have also been reports suggesting the involvement of SLIMB in TIM degradation [27].