
Figure 2. Effects of alanine substitutions in GFPΔ27BofA on inhibition of Pro-σK(1–127) cleavage in Escherichia coli.
pET Quartet plasmids were used to produce Pro-σK(1–127), cytTM-SpoIVFB, SpoIVFA, and GFPΔ27BofA (lane 1, pSO40) or Ala-substituted GFPΔ27BofA (lanes 2–20, pSO44-pSO58 and pSO60-pSO63). Samples collected after 2 hr of IPTG induction were subjected to immunoblot analysis with SpoIVFA, GFP, and penta-His antibodies. Single (*) and double (**) stars are explained in the Figure 1B legend, as is the graph.
Figure 2—source data 1. Immunoblot images (raw and annotated) and quantification of cleavage assays.
elife-74275-fig2-data1.zip (16.8MB, zip)

Figure 2—figure supplement 1. Sequence alignments of BofA orthologs to determine conserved residues.

(A) Sequence alignment of Bacillus subtilis BofA with 69 orthologs. Five residues (gray: M1, G51, I60, G69, and L71) are at least 70% conserved and nine residues (black: G40, L44, N48, N61, T64, G72, P74, G75, and L79) are at least 90% conserved, all within transmembrane segment 2 (TMS2) and the C-terminal region, except M1. (B) Sequence alignment of B. subtilis BofA with 30 orthologs that contain spoIVFA in their genome. In comparison to (A), four additional residues in predicted TMS2 and the C-terminal region were deemed of interest for Ala substitutions (H57, I82, and I86 are at least 70% conserved, and P59 is at least 90% conserved). A77 and A78 are also at least 70% conserved, but ineligible for Ala substitutions. V45 of B. subtilis does not match F, which is at least 70% conserved at the corresponding position of orthologs.
Figure 2—figure supplement 2. GFPΔ27BofA variants accumulate normally in Escherichia coli in the absence of the other Bacillus subtilis proteins.
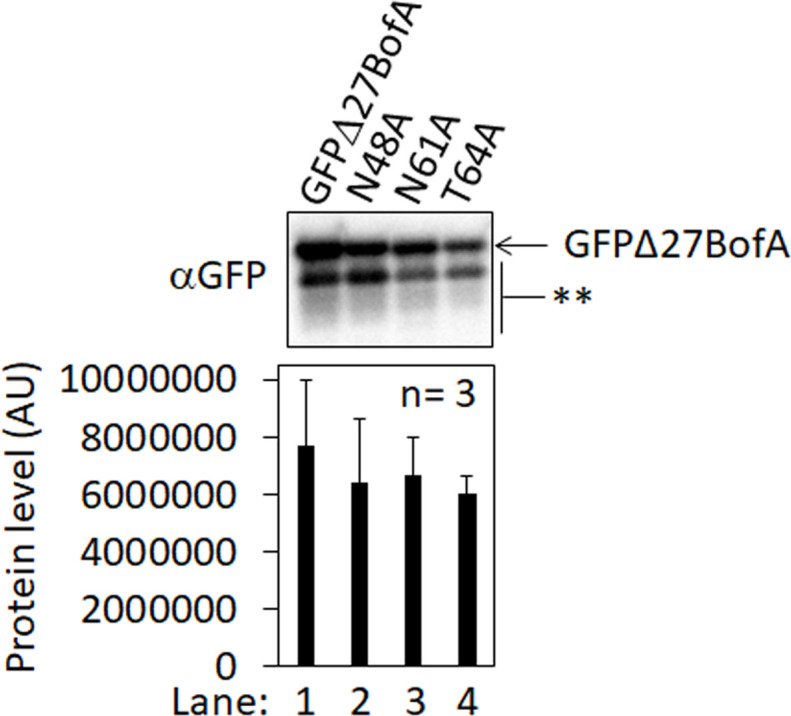
Plasmids were used to produce GFPΔ27BofA (pZR62, lane 1) or Ala-substituted GFPΔ27BofA (lanes 2–4, pSO332–pSO334). Samples collected after 2 hr of IPTG induction were subjected to immunoblot analysis with GFP antibodies. Double stars (**) indicate breakdown species of GFPΔ27BofA and its variants, which were not quantified. The graph shows the quantification of the GFPΔ27BofA and variant protein levels in arbitrary units (AUs) for three biological replicates. Error bars, 1 standard deviation. The protein levels of variants were indistinguishable from GFPΔ27BofA based on Student’s two-tailed t-tests (p>0.46).
Figure 2—figure supplement 2—source data 1. Immunoblot images (raw and annotated) and quantification of protein levels.
elife-74275-fig2-figsupp2-data1.zip (1.1MB, zip)
Figure 2—figure supplement 3. BofA C-terminal residues and residues preceding predicted TMS2 contribute to inhibition of SpoIVFB in Escherichia coli.

(A) Cleavage assays examining the effects of a GFPΔ27BofA truncation and a triple-Ala substitution for the last three residues of GFPΔ27BofA. pET Quartet plasmids were used to produce Pro-σK(1–127), cytTM-SpoIVFB, SpoIVFA, and GFPΔ27BofA from pSO40 as a control (lane 1), GFPΔ27BofA lacking the last three residues (Δ85–87) from pSO43 (lane 2), or GFPΔ27BofA with a triple-Ala substitution for the last three residues from pSO67. Samples collected after 2 hr of IPTG induction were subjected to immunoblot analysis, and the graph shows quantification of the cleavage ratio, as explained in the Figure 1B legend. For comparison, lanes 3–5 show data from Figure 2 for single-Ala substitutions in GFPΔ27BofA. The triple-Ala variant increased the cleavage ratio (lane 6), as did the variant lacking the three residues (lane 2). Both variants accumulated normally, but less SpoIVFA accumulated compared to the controls, indicating that residues near the C-terminal end of GFPΔ27BofA affect the synthesis and/or stability of SpoIVFA, and contribute to inhibition of cytTM-SpoIVFB in E. coli. (B) GFPΔ27BofA lacks TMS1 and all but nine residues preceding predicted TMS2 (Rudner and Losick, 2002). GFPΔ36BofA additionally lacks the nine residues. Cleavage assays were used to compare inhibition by GFPΔ27BofA, GFPΔ36BofA, or GFPΔ36BofA with a nine-residue glycine/serine (G/S) linker added between GFP and Δ36BofA. pET Quartet plasmids were used to produce Pro-σK(1–127), cytTM-SpoIVFB, SpoIVFA, and GFPΔ27BofA from pSO40 as a control lane 1, same data as in (A), GFPΔ36BofA from pSO42 (lane 2), or GFPΔ36BofA with the nine-residue G/S linker from pSO69 (lane 3). Samples were subjected to immunoblot analysis and quantification as in (A). Samples containing GFPΔ36BofA have a much greater cleavage ratio than samples containing GFPΔ27BofA. Since all four proteins accumulated well in both cases, the nine residues appeared to contribute to the inhibitory function of GFPΔ27BofA. Replacement of the nine residues with the G/S linker decreased the cleavage ratio, based on a Student’s two-tailed t-test (p value is indicated), suggesting that moving the GFP tag away from the membrane restored inhibitory function almost completely.
Figure 2—figure supplement 3—source data 1. Immunoblot images (raw and annotated) and quantification of cleavage assays.
elife-74275-fig2-figsupp3-data1.zip (8.6MB, zip)