
Fig. 6 |. BSL requires BIN2 and YDA to regulate stomatal development in Arabidopsis.
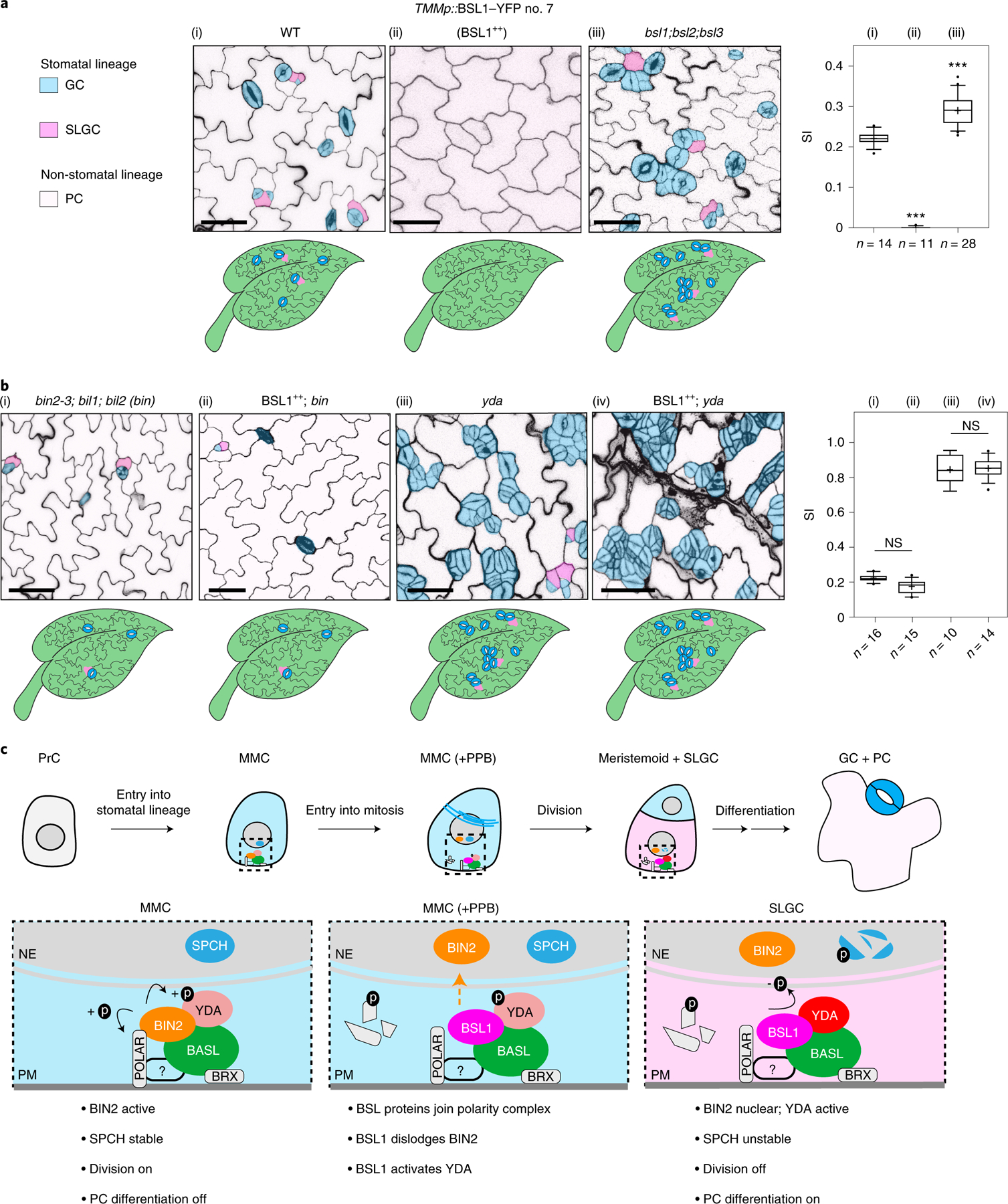
a, BSL proteins function to suppress stomatal production. Left: confocal images (converted to black and white) showing 5-day adaxial cotyledon epidermis of the designated genotypes. BSL1++, overexpression of BSL1 by the stomatal lineage TMM promoter. Stomatal lineage cells were manually traced and highlighted by different shadings. Blue, stomatal GCs; pink, SLGCs; light pink, PCs. Right: quantification of SI (number of stomata relative to the number of total cells). Box plots show the first and third quartiles, median (line) and mean (cross). n, number of cotyledons. One-way ANOVA with Tukey’s post-hoc test were used to compare with the WT. ***P < 0.0001. b, BSL requires BIN2 and YDA to regulate stomatal development. Confocal images show stomatal phenotypes when overexpression of BSL1 (BSL1++) was introduced in the loss-of-function bin (left) or yda (right) mutants, respectively. Right, quantification of SI of the designated genotypes. Box plots show the first and third quartiles, median (line) and mean (cross). n, number of cotyledons. Two-tailed Student’s t-tests were used to compare with the respective mutant background. Scale bar, 40 μm (a and b). The cartoons in a and b depict the corresponding stomatal phenotype shown above. Blue, stomatal GCs; pink, stomatal lineage cells; puzzle shapes, PCs. c, Working model whereby BSL proteins function as a spatiotemporal molecular switch enabling stomatal ACD. In the MMC, the high cell division potential is maintained by the BIN2 GSK3-like kinases that associate with the BASL polarity complex via POLAR to be enriched at the cell membrane19, where BIN2 suppresses the MAPKK kinase YDA and MAPK signalling22. Therefore, SPCH activity is maintained at high levels to promote cell division. In this study, we identified the association of the BSL proteins with the BASL polarity complex, and the polarization of the founding member BSL1 coincides with the formation of the PPB (blue lines in the MMC) at the entry of MMC to mitosis. The participation of BSL proteins in the polarity complex may directly or indirectly dissociate BIN2 from the plasma membrane, releasing its inhibition on YDA MAPK signalling. Polarized BSL1 is inherited by the SLGC, in which its phosphatase function could directly activate YDA MAPK signalling, leading to strong suppression of SPCH and PC differentiation. Thus, we propose that polarized BSL1, by jointly regulating BIN2 GSK kinase and YDA MAPK activities, functions as a spatiotemporal molecular switch to establish a kinase-based signalling asymmetry that enables cell-fate asymmetry in the two daughter cells. See Discussion for more details. Graphics on top show the progressive stages of stomatal ACD in Arabidopsis. Dotted rectangles represent the regions enlarged at the bottom containing the polarity complex in the MMC (left), PPB-containing MMC (middle) and SLGC (right). Light blue, stomatal fate; pink, non-stomatal fate. NE, nuclear envelope; PM, plasma membrane. The question mark indicates unidentified regulator(s) for POLAR to associate with the polarity complex.