
Extended Data Fig. 1 |. Genetic Pathways Involving BSL Proteins in Arabidopsis.
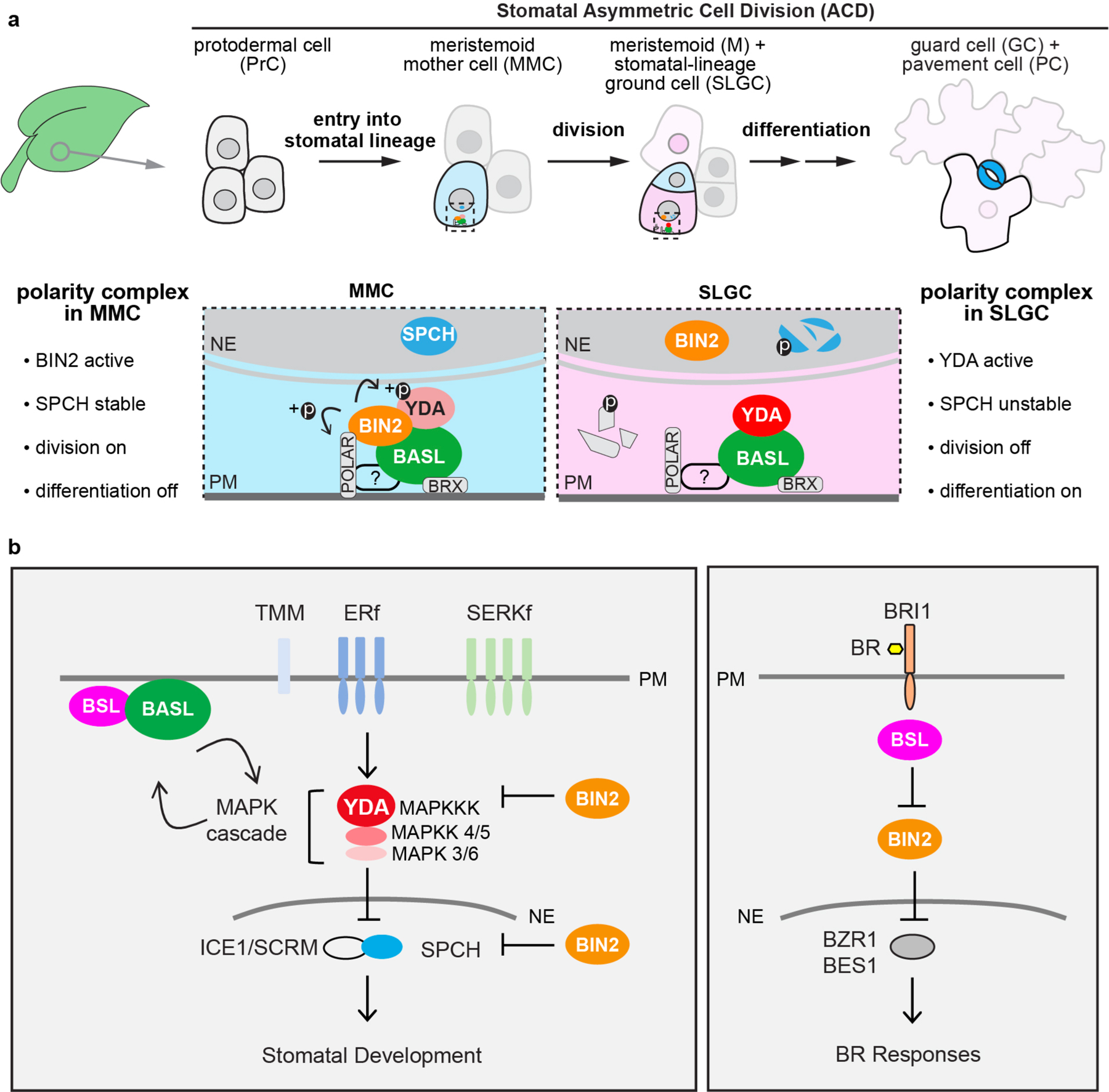
a, Compositional and functional changes of the polarity complex before and after a stomatal ACD. In a young leaf, the initiation of the stomatal lineage cell (MMCs), is driven by the expression of the transcription factor SPEECHLESS (SPCH)11. The plant-specific protein BASL is polarized before, during and after a stomatal ACD13. Multiple components in the BASL polarity complex have been identified to regulate cellular events in stomatal ACD. The BRX proteins interact with BASL to attach the polarity complex to the plasma membrane15. The POLAR proteins associate with the polarity complex and require BASL to become polarized14. It is known that POLAR recruits the BIN2 GSK3-like kinases to the polarity complex in the MMC, where BIN2 phosphorylates and inhibits the MAPKKK YDA, leading to alleviated MAPK-mediated suppression of SPCH, thereby the MMC undergoes cell division19. It is also known that, BASL interacts with YDA to locally concentrate MAPK signaling, thus SPCH is suppressed in the large daughter cell SLGC that inherits the polarity complex, thereby the SLGC after an ACD undergoes differentiation to become a pavement cell18,35. Thus, a successful stomatal ACD necessitates the changes of the key regulators, that is BIN2 to be preferentially membrane-localized before ACD but nucleus-partitioned after ACD, whereas YDA to be suppressed before ACD but activated after ACD. Light blue, stomatal fate; dark blue, stomatal guard cells; pink, non-stomatal fate; light pink, pavement cell. Fading cells, PrCs not converted to MMC become non-stomatal pavement cells. Dotted rectangle, regions enlarged in bottom, containing protein components of the polarity complex. Bottom, enlarged view of polarity protein complexes required for the progenitor cell (MMC, light blue) and the daughter cell (SLGC, pink), respectively, in stomatal ACD. ?, unidentified regulator/s for POLAR to associate with the polarity complex. Fragmented molecules (SPCH and POLAR) indicate proteins undergo degradation. b, Schematics depicting simplified signal transduction pathways in stomatal development (left) and Brassinosteroid (BR) signaling (right). The models are mainly based on Arabidopsis research. In stomatal development, the receptor-like kinases/protein (TMM, TOO MANY MOUTHS; ERf, ERECTA family; SERKf, SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE family)27,30,31 transduce signals from the EPF peptide ligands (not specified here) to activate the YODA (YDA) MAPK signaling that suppresses the transcription factors (SPCH, SPEECHLESS; ICE1/SCRM, SCREAM) and stomatal development17,20,56. The BIN2 GSK-3 like kinases regulate stomatal development by inhibiting YDA and SPCH21,22. The polarity protein BASL interacts with YDA and forms a positive feedback regulation with the MAPK cascade to regulate stomatal ACD18. In this study, we identified the BSL protein phosphatases as BASL partners in the polarity complex. In BR signaling, the hormone (BR) is perceived by the receptor-like kinase (BRI1, BRASSINOSTEROID INSENSITIVE 1)57 that triggers cytoplasmic signal transduction mediated by the BSL protein phosphatases23 and the BIN2 GSK3-like kinases58 to ultimately active the expression of the transcription factors (BZR1, BES1)59,60 for plant responses. Both pathways involve BSL Ser/Thr protein phosphatases and the GSK3-like BIN2 kinase. Block lines indicate negative regulation and arrows indicate positive regulation. PM, plasma membrane; NE, nuclear envelope.