
FIGURE 5.
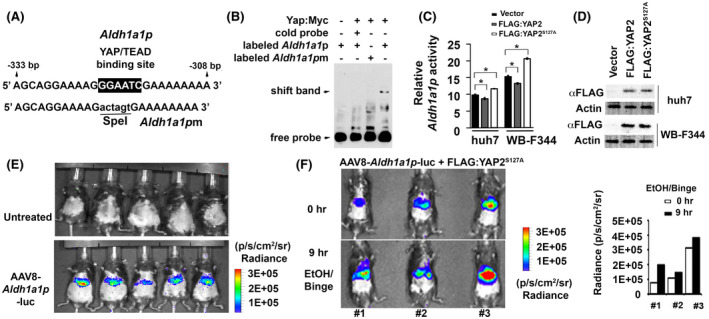
YAP can transcriptionally regulate Aldh1a1 promoter activity in vitro and in vivo. (A) Illustrates sequences flanking a putative YAP/TEAD binding site in wild type promoter (Aldh1a1p) while mutated fragments without this site (Aldh1a1pm) are used to determine binding specificity. (B) Complexes between a murine Yap protein that was fused with Myc epitope at its C terminal region (Yap:Myc) and biotinylated Aldh1a1p fragments were formed, whereas this binding was disrupted in presence of excessive cold or mutant probes in gel shift assays. (C) Aldh1a1 promoter activity as indicated by ratio between Gluc and SEAP could be induced by the FLAG:YAP2S127A mutant, but was inhibited by FLAG:YAP2 protein in Huh7 and WB‐F344 cells. Data were expressed as means ± SD in triplicate experiments. *p < .05. (D) Immunoblotting detected expression of FLAG:YAP2 and FLAG:YAP2S127A proteins in tested cells. (E) Bioluminescence showed Aldh1a1‐promoter driven luciferase activity in AAV8 virus that was transduced into mouse livers through tail vein injection, whereas no signal was seen in untreated control animals. (F) Bioluminescence detected induction of Aldh1a1p activity at 9 h compared to 0 h after binge in ethanol‐fed mouse livers that were transfected with the FLAG:YAP2S127A mutant. Graphs were quantification that determined the Aldh1a1p driven luciferase activity based on bioluminescent signals (Radiance, p/s/cm2/sr)