

Abstract
By collecting data continuously over 24 hours, accelerometers and other wearable devices can provide novel insights into circadian rhythms and their relationship to human health. Existing approaches for analyzing diurnal patterns using these data, including the cosinor model and functional principal components analysis, have revealed and quantified population-level diurnal patterns, but considerable subject-level variability remained uncaptured in features such as wake/sleep times and activity intensity. This remaining informative variability could provide a better understanding of chronotypes, or behavioral manifestations of one’s underlying 24-hour rhythm. Curve registration, or alignment, is a technique in functional data analysis that separates “vertical” variability in activity intensity from “horizontal” variability in time-dependent markers like wake and sleep times; this data-driven approach is well-suited to studying chronotypes using accelerometer data. We develop a parametric registration framework for 24-hour accelerometric rest-activity profiles represented as dichotomized into epoch-level states of activity or rest. Specifically, we estimate subject-specific piecewise linear time-warping functions parametrized with a small set of parameters. We apply this method to data from the Baltimore Longitudinal Study of Aging and illustrate how estimated parameters give a more flexible quantification of chronotypes compared to traditional approaches.
Keywords: accelerometer, data-driven chronotypes, diurnal registration, cosinor model, functional principal components analysis
1. Introduction
A person’s circadian rhythm is driven by the internal clock that dictates his or her daily physiological cycles, including sleep-wake times, hormone release, core body temperature, blood sugar regulation, and other components of the homeostatic system, all of which influence how one distributes his or her daily activity (Zee et al., 2013). Although circadian rhythms can be influenced by social and solar clocks, they are endogenous and determined largely genetically (Roenneberg et al., 2003). Chronotypes, or phenotypes of circadian rhythms that can be used to identify population subgroups and classify individuals, have been linked to numerous health outcomes. For example, some individuals wake earlier in the day (i.e., a “morningness chronotype”) while others stay awake later into the night (i.e., a “eveningness chronotype”). Those with an eveningness chronotype also tend to be at greater risk for some mental health disorders and general health problems such as hypertension, asthma, and type 2 diabetes (Partonen, 2015). Chronotypes have typically been defined on a single scale ranging from morningness to eveningness (Dunlap et al., 2004; Partonen, 2015). Methods have been developed to estimate chronotype along this scale as a continuous or dichotomous variable, using measurements obtained from either body temperature and hormone levels (Dunlap et al., 2004), self-reported wake and sleep times via self-report (Horne & Ostberg, 1976; Roenneberg et al., 2007) or actigraphy (Urbanek et al., 2018).
Despite the current emphasis on a single morningness to eveningness scale to describe chronotypes, there may be more nuanced phenotypes characterizing one’s pattern of energy over the course of a day, independent of sleep and wake time preferences. For example, a recent study revealed that measures of sleep quality, sleep duration, and diurnal inactivity (e.g. nap times or wakeful rest) showed very little correlation with self-reported chronotype on a scale from “definitely a ‘morning’ person” to “definitely an ‘evening’ person”, suggesting that these other sleep features provide new information beyond the established chronotypes (Jones et al., 2019). The sleep and diurnal features examined were extracted from accelerometers, which give rich data for studying daily activity patterns by collecting epoch-by-epoch (often minute-by-minute) data on activity intensity over 24 hours. Accelerometers are a natural mechanism for tracking diurnal patterns in order to detect underlying circadian rhythms. Novel data-driven techniques are now capable to leverage the rich information collected by accelerometers to confirm the existence of morningness and eveningness chronotypes, to identify more subtle differences in chronotypes, and to illustrate how these are manifested in various epidemiological cohorts and clinical sub-groups.
We introduce a new accelerometer data-driven approach, based on curve registration (Wrobel et al., 2019), for understanding circadian rhythms. Curve registration uses tools from functional data analysis, a statistical subfield that focuses on the kind of intensive longitudinal data made possible by wearable devices and other technologies (Ramsay & Silverman, 2002, 2005). Our method separates and analyzes both 1) “horizontal” variability in time-dependent markers like wake and sleep times, and 2) “vertical” variability in activity intensity; by doing so, the method gives distinct insights into underlying circadian rhythms. The remainder of this paper is organized as follows. In Section 1.1, we provide a review of existing approaches for using accelerometer data to study circadian rhythms. In Section 2, we describe the study methods, including our proposed registration method and motivating data from the Baltimore Longitudinal Study of Aging (BLSA). In Section 3, we use the registration method to analyze accelerometer data from the BLSA and compare results to those of existing methods. We conclude with a discussion in Section 4.
1.1. Review of existing approaches
Circadian rhythms can be studied using a range of data collection instruments. The temporality provided by diurnally measured core body temperature, melatonin, and cortisol is considered to be the most accurate way to determine chronotype (Dunlap et al., 2004; Melo et al., 2017), though challenging to measure. Self-reports such as the Horne-Ostberg Morningness-Eveningness Questionnaire (MEQ) (Horne & Ostberg, 1976) and the Munich ChronoType Questionaire (MCTQ) (Roenneberg et al., 2003, 2007) are also instruments for collecting chronotype information. The MCTQ for example asks questions about sleep routines on work and work-free days including questions about when respondents go to bed and when they get out of bed. The midpoint of the sleep interval as determined from self-reports can be used to describe chronotype on a morningness-eveningness scale (Roenneberg et al., 2003, 2007).
Below we highlight methods for evaluating chronotype that involve the collection of minute-by-minute accelerometer data. We categorize accelerometer-based methods into two distinct classes: landmark methods, which extract predefined features such as sleep and wake times, and methods that use the full 24-hour activity profile in an attempt to quantify the complexity of diurnal patterns.
Landmark methods define chronotypes based on one or two summary measures of a subset of one’s 24-hour period. Some of these methods identify a fixed-length interval, including the 5 least active consecutive hours and the 10 most active consecutive hours, often abbreviated L5 and M10, respectively, in the chronotype literature. The timing of such intervals can indicate morningness versus eveningness preferences (Witting et al., 1990). Other landmark methods identify intervals whose length varies across subjects, such as sleep or daytime intervals (Gershon et al., 2018; Kaufmann et al., 2018; Urbanek et al., 2018). One example is the sleep period time window (SPT-window) approach of van Hees et al. (2015, 2018). The SPT-window, developed for 3-axial raw accelerometer data, is a heuristic approach that entails searching within the 12-hour period centered around the L5 midpoint for periods of “sustained inactivity,” defined by absence of change in arm angle. The SPT-window then stretches from the start of the first period of sustained inactivity to the end of the last period within that 12-hour span. For sleep and daytime intervals, both the duration and the timing of the interval’s midpoint can be assessed, although the midpoint alone is often used to define chronotypes on either a continuous scale (Urbanek et al., 2018) or dichotomously by establishing some cut-off for the midpoint (Gershon et al., 2018; Kaufmann et al., 2018).
All landmark methods suffer from the same drawback: they condense activity data to produce a single, pre-defined aggregate measure (or a few such measures) within long time windows. This can eliminate subtle, but important and relevant differences in daily accelerometry profiles which may help to differentiate, for example, someone who is an early riser but is sedentary in the afternoon from someone who is an early riser and remains continuously active throughout the day. Moreover, only the data inside the landmark interval of interest is summarized, rather than the full 24-hour rest-activity profile. Lastly, for those methods that use only the timing of an interval’s midpoint to define chronotypes, duration of activity and sleep is not factored into the chronotype at all; someone who sleeps from 10PM to 6AM has the same sleep interval midpoint as someone who sleeps from 11PM to 5AM. Therefore, without separate adjustment for sleep duration, landmark methods may incorrectly classify a broad range of circadian rhythm patterns into a single chronotype.
Other methods utilize 24-hour rest-activity profile, leveraging the complex information and extracting more intricate features of circadian rhythms. The cosinor method captures both the timing and intensity of one’s daily active period in a single three-parameter model. It does so using a parametric regression model with clinically interpretable parameters, including the MESOR (midline estimating statistic of rhythm) or model-based mean intensity, the amplitude or peak intensity, and the acrophase or timing of the peak intensity (Halberg et al., 1967; Marler et al., 2006). The acrophase parameter in particular has been shown to strongly correlate with the MEQ (coefficient of determination = 0.7) (Roveda et al., 2017). The cosinor method has been widely used to study circadian rhythms (Cornelissen, 2014; Refinetti et al., 2007). However, a major limitation of this method is that it assumes all activity curves conform to a cosine shape: this fit may be too restrictive to accurately represent the circadian rhythm profile for many subjects.
Like the cosinor approach, functional principal components analysis (FPCA) models epoch-by-epoch accelerometry data; however, FPCA is a data-driven approach that does not impose any pre-defined parametric assumptions about the shape of daily rest-activity rhythms. Each person’s 24-hour rest-activity profile is represented via a combination of common population-level patterns (principal components) that capture most of the variability across subjects representing the population. Principal component (PC) scores for a given subject each indicate the degree to which these patterns are present for that subject and tell a piece of the circadian rhythm puzzle: whether he or she has a generally more active day, whether he or she wakes up early or late, how long his or her day is, and other specific and independent aspects of the circadian rhythm. FPCA has recently gained traction within the sleep research community (Gershon et al., 2016; Zeitzer et al., 2013), in part due to the appeal of its flexibility. However, 24-hour rest-activity profiles have two distinct sources of variability: horizontal variability, which reflects the timing of features like wake and sleep times and peaks of activity, and vertical variability, which reflects the intensity of activity. FPCA does not separate these distinct sources of variability, and the patterns that are identified can conflate the two sources in ways that could mask the true behavioral differences between people.
The registration method introduced in this paper retains the strengths of existing approaches while addressing their limitations. By using the full 24-hour rest-activity profile, registration can identify differences in landmarks such as sleep and wake times, but also provides more detailed information than landmark methods. In contrast to the cosinor approach, registration does not place a restrictive shape on the data, but still produces interpretable and clinically meaningful parameters. Registration builds on FPCA as a standalone method by separating horizontal variability (timing of activity) from vertical variability (intensity of activity), which results in more easily interpretable PCs. We will show that the registration method, by explicitly separating the sources of variability, reveals new insights into accelerometry-estimated chronotypes and circadian rhythms.
2. Methods
2.1. Our proposed registration method
2.1.1. Overview
Our approach brings two new ideas to the study of circadian rhythms. First, we dichotomize the minute-level activity counts into “active” or “resting” states rather than directly analyzing minute-level (or other epoch-level) activity counts like the methods discussed in the previous section. When activity counts are analyzed directly, the periods with high-intensity exercise or other strenuous activities may heavily influence the shape of one’s activity profile and consequently our inference about one’s chronotype. Another limitation of existing approaches is that they provide results that depend on the definition or scale of activity count that is often device- and manufacturer-specific. We expect our approach to be invariant to the scale of activity count because we dichotomize minute-level activity-counts with the objective of detecting patterns in the probability of active state over time, which we term “activity probability profiles”. Note that we use “resting” to refer both to “sedentary” (awake, but not active) and sleep periods.
Second, we introduce registration in the context of accelerometry-estimated circadian rhythms. The goal of our method is to align subjects’ activity probability profiles – smooth, underlying curves that quantify the probability that a subject is active at each time of the day – based on shared features such as wake and sleep times and peaks in activity probability. We accomplish this by separating the variability across activity probability profiles into the horizonal and vertical sources. Figures 1 and 2 illustrate this process. In the left panel of Figure 1, each curve represents a different subject’s activity probability profile over a 24-hour period, with one subject highlighted in red. Differences across subjects in the timing of common features such as sleep and wake times make it difficult to identify any shared daily patterns. In the right panel of Figure 1, we have used the proposed registration approach to align these curves and a clearer shared pattern is revealed, showing a morning peak, a midday dip in activity, and an afternoon peak; nearly all variability in the right panel of Figure 1 is vertical variability. The transformation of curves in the left panel to curves in the right panel is accomplished through subject-specific (curve-specific) time-warping functions shown in the middle panel of Figure 1, which capture the horizontal variability.
Figure 1.

Activity probability profiles pre-registration (left), warping functions (middle), and activity probability profiles post-registration (right). In each panel, each curve represents data for a particular subject. Data from one subject is highlighted in red across all panels.
Figure 2.

Example of a warping function for one subject mapping chronological time to registered time. The left panel demonstrates where certain points throughout his chronological day fall on his registered timeline. The right panel displays the corresponding 2-knot piecewise linear warping function.
Warping functions map observed “chronological” time to a latent “registered” time domain on which all profiles are aligned. Figure 2 illustrates the effect of the warping function for the subject highlighted in Figure 1. In the left panel of Figure 2, this subject’s window of activity between 3AM and 9AM in chronological time (top) becomes stretched, or slowed down in registered time (bottom), so that it covers nearly half the day. In other words, the activity profile that this subject experienced in the first 6 hours of their morning, most people take until just before 3PM to complete. Conversely, the 15-hour window from 9AM to nearly midnight in chronological time becomes compressed or sped up in registered time, so that it only accounts for what would be a typical late afternoon and evening. After midnight time is similar between chronological and registered timelines, suggesting this subject has an average bedtime relative to the study population. The shape of the warping function in the right panel of Figure 2 reflects this exact mapping from chronological time (x-axis) to registered time (y-axis). Note that our registration framework assumes two time points (knots) that split the 24-hour period into three different periods with different linear dependences (slopes or angles of the linear lines) between the chronological and registered times. Each subject’s set of parameters, i.e., the two knots and three slopes, describe how the timing of his or her activity profile features differ from the population mean. We expand on this in more technical detail below.
2.1.2. Technical details
Registration accomplishes the separation of horizontal and vertical variability using an iterative two-step approach, cycling between estimating warping functions to capture the horizontal source of variability, and applying FPCA to understand vertical variability in the post-warped/registered patterns. We will begin with the mathematical representation of this two-step iterative process and then provide a more heuristic breakdown of each step. The following notation draws from the statistical subfield of functional data analysis, which provides a useful conceptual framework for intensive longitudinal data (e.g., minute-by-minute rest/active states over 24 hours). Let represent chronological time and let represent the subject’s activity probability profile over chronological time. Warping functions map observed chronological time to registered time . The resulting unregistered and registered activity probability profiles can be written as and , respectively; this notation implies activity probability profiles observed over continuous time, although in practice we observe these profiles at discrete times (for example, at minute level). The warping functions align the observed activity probability profiles to subject-specific mean templates , which are estimated using a binary form of FPCA. Notationally, we combine warping functions with FPCA through the following:
| (1) |
where are the PC functions, are subject-specific PC scores, and is the population mean function. In the first step of Equation (1), we estimate the warping functions while holding the mean templates fixed. In the second step, we estimate the mean templates using FPCA on the registered data while holding fixed. We iterate through these two steps until convergence. Below we explain these two steps in greater detail; a full technical overview of registration can be found in (Wrobel et al., 2019).
Registration step 1. Warping functions.
In the first step of Equation (1), we estimate subject-specific warping functions to map chronological time to registered time so that the resulting registered activity probability profiles are aligned to the most recent mean templates . The warping functions can take many forms; in this paper, we define a 2-knot piecewise linear function that is novel in the literature, elegant in its simplicity and interpretability, and flexible enough to accurately register daily activity profiles. As demonstrated in Figures 1 and 2, the function for a given subject consists of a series of three slopes connected at two knot locations along the chronological timeline:
| (2) |
For simplicity, we rescale the 24-hour period so that time is defined on We place additional constraints on the warping functions to ensure that they are always increasing (i.e., we never warp time backwards) and that times 0 (the beginning of the 24-hour period) and 1 (the end of the 24-period) always map to 0 and 1, respectively. The three slopes and two knots that make up each subject’s warping function convey information about his or her diurnal pattern relative to the population.
Registration step 2. FPCA.
In the second step of Equation (1), we use FPCA to estimate subject-specific mean templates , using the most recent warping functions to define registered time . In other words, we perform FPCA on the newly registered activity probability profiles . Because we propose to dichotomize the activity counts, we use the extension of FPCA that applies to binary functional data (Goldsmith et al., 2015; van der Linde, 2008). Binary FPCA estimates a functional template of the probability of activity over registered time for each subject, which reflects the population mean behavior while allowing for subject-specific deviation. Specifically, at each iteration of this step we estimate a population-level functional mean population-level functional PCs , and subject-specific scores which indicate how much each component contributes to the subject-specific activity probability template. These population- and subject-level parameters are combined to provide an updated estimate of as defined in Equation (1).
Iterate between the warping function and FPCA steps.
Registration iterates between these two steps of Equation (1) until the activity probability curves are temporally aligned across subjects. After registration, we have separated the original activity probability profiles into warping functions and aligned activity probability profiles, which can each be used to understand different aspects of individual variation in circadian rhythms. From the warping functions, subject-specific slopes reveal information about the increasing/decreasing nature of subject-specific time with respect to the registered time and the two times of the day when the linear slopes defining the subject-specific alignment change. From the aligned activity profiles, we can learn about both population-level and subject-specific activity patterns. The estimated population mean activity profile and PC functions describe a typical 24-hour activity probability profile and the major sources of variability across subjects, respectively. The subject-specific PC scores indicate individual deviations from the population mean profile after removing any variability in the timing of landmark features. All of these pieces of information contain useful insights into heterogenous complex nature of chronotypes across subjects. The registration procedure is implemented in the R package “registr” (Wrobel et al., 2020).
2.2. Motivating data
Our motivating data come from the Baltimore Longitudinal Study of Aging (BLSA), a prospective observational study collecting health, cognitive, and physical performance evaluations of initially healthy participants every 1–4 years for life. Data were collected by the Intramural Research Program of the National Institute on Aging and the protocol for this secondary analysis was approved by the Institutional Review Board of Columbia University. All subjects provided written informed consent to participate in the study. Actiheart activity monitors (Schrack et al., 2014) were worn by the participants for up to seven consecutive days. We illustrate our approach by analyzing the first available Tuesday from each subject, chosen to represent typical weekday activity without the after-weekend inertia typically observed on Mondays (Urbanek et al., 2018). Twenty-four-hour activity periods began at 3AM of the specified day and ended at 2:59AM of the following day. Minute-level activity counts were dichotomized into active (activity count > 10) or resting (activity count ≤ 10) state, using a threshold that was previously established for the BLSA Actiheart data (Schrack et al., 2018, 2019; Wanigatunga et al., 2019). This value was specifically selected for this device and study population by first separating patients’ minute-level activity into sedentary, light, moderate, and vigorous activity based on heart rate reserve (HRR) levels measured using continuous recordings from a heart rate monitor, and then examining distributions of activity counts within those categories. Note that this threshold is specific to the BLSA Actiheart data; other devices or populations may require different cut points.
3. Results
3.1. Registration application
After data processing, 492 subjects were analyzed. The median age was 71 years (interquartile range: 62 to 80), and 54% were male. We applied the registration method to these 24-hour rest/active binary profiles, using two PCs for the binary FPCA step. The left, middle, and right panels of Figure 1 display unregistered activity probability profiles obtained by smoothing minute-level rest/active binary curves, estimated warping functions, and registered activity probability profiles, respectively. At the population level, the registered curves in the right panel reveal a common trend across subjects, which includes a morning activity peak, a midday dip during which subjects are in a more restful state, and a late afternoon/evening activity peak. The horizontal variability in activity data is captured in the warping functions and remaining vertical variability in activity probability profiles is captured through the FPCA decomposition of the registered data; these are both interpretable in the context of chronobiology.
The warping functions are parameterized by three subject-specific slopes given in Equation (2), each of which reveal different information about morningness and eveningness. A steep first line segment (slope > 1) indicates that a subject’s activity during the late night and morning gets moved forward in time to match the population pattern — that is, his or her day started earlier in chronological time than the average BLSA subject. An extremely steep first line segment suggests an “early bird”, similar to the example subject from Figures 1 and 2. On the contrary, a warping function with a flatter first line segment (slope < 1) indicates that one’s day started later in chronological time. We can apply analogous interpretations to the third line segment of the warping function, which generally takes place during evening and nighttime hours: extremely steep third line segments indicate “night owls”, while flatter third line segments represent subjects who end their active day earlier than most. When a subject has a warping function close to the identity line (i.e., all three slopes near 1), this suggests that his or her activity probability profile in chronological time was in line with the population mean and therefore needed little to no warping to map to the registered time domain.
The registered activity probability profiles are parameterized by the subject-specific scores from the two PCs estimated using binary FPCA. Figure 3 shows the population mean activity probability profile in black plus (blue) or minus (red) one standard deviation of the first (left panel) or second (right panel) PC score. Together these panels show the patterns identified by FPCA for these data. The first PC captures variability across subjects in terms of overall activity probability. The second PC captures variability across subjects in terms of which activity peak (morning or evening) is more dominant. This should not be confused with morningness vs. eveningness chronotypes, which are a reflection of sleep timing (and are better captured by warping function parameters). Some remaining variability in wake times that was not captured in the warping functions is indeed evident in this second PC, as well as some potential restlessness at the end of the activity profile. However, the second PC largely indicates whether a subject is most consistently active immediately upon waking or later in the day.
Figure 3.

Population mean activity probability profile from registration’s binary FPCA step (black lines), plus or minus some variability (red and blue lines, respectively). The left panel demonstrates the mean +/− 1 standard deviation in the first principal component, and the right panel demonstrates the mean +/− 1 standard deviation in the second component.
Pairwise correlations between the five registration parameters (three warping function slopes and two PC scores) reveals low correlation among the first and third slopes and the PC scores (absolute value of Pearson correlation coefficients ≤ 0.12), but higher correlation between the second slope and the other two slopes (absolute value of Pearson correlation coefficients ≥ 0.44, Supplementary Figure S1). As such, in the remainder of this paper we focus our interpretations on the two PC scores and the first and third slopes. Each of the axes (parameters) in this four-dimensional space has a clear interpretation and is not strongly correlated with the other axes.
This continuous four-dimensional parametric space is well-suited to describe complex between-subject differences in accelerometry-estimated circadian rhythms, going beyond the continuous one-dimensional morningness-eveningness scale. Extreme values on these four dimensions may be interesting and may yield new, more nuanced description of the complexity of observed chronotypes. For example, extreme values of the warping function slopes classify subjects by their wake and sleep times, leading to chronotypes of “morning lark” vs. “anti-lark” (early vs. late wake time) and “night owl” vs. “anti-owl” (late vs. early sleep time). Extreme values of the first PC score reveal potential chronotypes of “penguin” vs. “hummingbird” (low vs. high overall activity probability); and extreme values of the second PC score reveal potential chronotypes of “rooster” vs. “roadrunner” (most active in the morning vs. afternoon). These potential chronotypes are summarized in Table 1, with corresponding simulated activity probability profiles displayed in Figure 4. In Figure 4, for chronotypes that are defined by warping function slopes, defining features of these chronotypes are most evident before registration, i.e., before the warping functions absorb horizontal variability or differences in sleep and wake times. Conversely, defining features of the PC-score-based chronotypes are more strongly evident after registration, once probability activity profiles have been aligned and all that remains is vertical variability.
Table 1.
Potential chronotypes defined by cut-off values of registration parameters.
| Registration parameter (for subject ) | Values | Proposed chronotype |
Defining feature |
|---|---|---|---|
| (1st slope of warping function) | High | Lark | Early wake time |
| Low | Anti-lark | Late wake time | |
| (3rd slope of warping function) | High | Owl | Late sleep time |
| Low | Anti-owl | Early sleep time | |
| (1st PC score) | High | Penguin | Low overall activity probability |
| Low | Hummingbird | High overall activity probability | |
| (2nd PC score) | High | Rooster | Most active during (registered) morning |
| Low | Roadrunner | Most active during (registered) afternoon |
Figure 4.

Simulated activity probability profiles from select chronotypes for demonstration. The top row compares a lark and an owl in terms of their profiles in chronological time (top left), warping functions, (top middle), and profiles in registered time (top right). In the bottom left, the profiles of a hummingbird and a penguin are compared in registered time. In the bottom right, the profiles of a roadrunner and a rooster are compared in registered time. These simulated profiles were generated by specifying warping function parameters and PC scores that meet each chronotype’s definition. We refer the reader to Table 1 for these definitions.
We emphasize that these chronotypes are not mutually exclusive, but rather describe four independent domains on which a subject can be typical or outlying. A clustering analysis validates our proposed chronotypes and illustrates that multi-dimensional approaches can uncover sophisticated subtypes of behavior. By applying k-means clustering to the four-dimensional registration parameter space, and considering a balance of average silhouette width and interpretability, we identified six clusters. The six subgroups identified by this unsupervised learning method have defining features that align with the chronotypes proposed in Table 1. The clusters are presented in Figure 5, with labels corresponding to their most distinguishing feature(s). Two clusters contained the larks and owls, whose deviations from the population mean activity probability profile were mainly temporal shifts or horizontal variability. The remaining four clusters grouped subjects by patterns of activity characterized by the PC scores. The ability to define chronotypes across all four dimensions of the registration model may lead to the identification of more sophisticated and distinct subgroups of circadian rhythm patterns compared to existing methods.
Figure 5.

Results from k-means clustering on the registration’s 4-dimensional parametric space. Top row shows data under chronological time, and the bottom row shows data under registered time. Columns contain distinct clusters, and cluster labels correspond to the chronotypes proposed in Table 1 and they are chosen to reflect each cluster’s most distinguishing features.
3.2. Comparison to existing methods
We compared our approach to three existing methods: i) the SPT-window landmark method, ii) the cosinor method, and iii) binary FPCA without registration.
3.2.1. Comparing registration with landmark times
For the SPT-window, we lacked the tri-axial data necessary to calculate arm angle, so we modified the definition of sustained inactivity to be a status of rest for a duration of more than 20 consecutive minutes. This 20-minute criterion was chosen post-hoc after visually inspecting its ability to identify approximate sleep and wake times. We estimated two SPT-windows: one during the night between Monday noon and Tuesday noon, and one during the night between Tuesday noon and Wednesday noon. The end of the former SPT-window defined the Tuesday wake time, while the start of the latter SPT-window defined the Tuesday sleep time. Using this approach, we identified wake and sleep times for 424 of the 492 subjects.
Figure 6 contains two panels of lasagna plots (Swihart et al., 2010), in which each row represents a different subject’s rest/activity status over 24 hours, with blue cells representing active minutes and white cells representing rest minutes. In each panel, wake and sleep times for each subject are highlighted in black and the daytime interval midpoint (DIM) for each subject is highlighted in red. Landmark times were first estimated on the chronological time scale (left); and then we evaluated where these landmarks mapped to in registered time (right). Before registration, we first take note that DIMs are not a strong indicator of morningness vs. eveningness; there is considerable overlap in DIM between those who wake later (near the top of the plot) and those who wake earlier (near the bottom of the plot). After registration, we see better alignment in sleep times and wake times as expected, although there is still some between-subject variability, particularly in sleep times. What is more concerning, however, is that the DIM does not consistently map to an intuitively meaningful feature of the registered activity probability profiles. For some patients, the DIM takes place during their midday dip or period of rest. For other patients, it takes place at the height of their morning peak, and still for others, it takes place toward the end of their evening activity peak. The DIM, which is simply calculated as the halfway point between wake and sleep, fails to define any single feature of one’s circadian rhythm.
Figure 6.
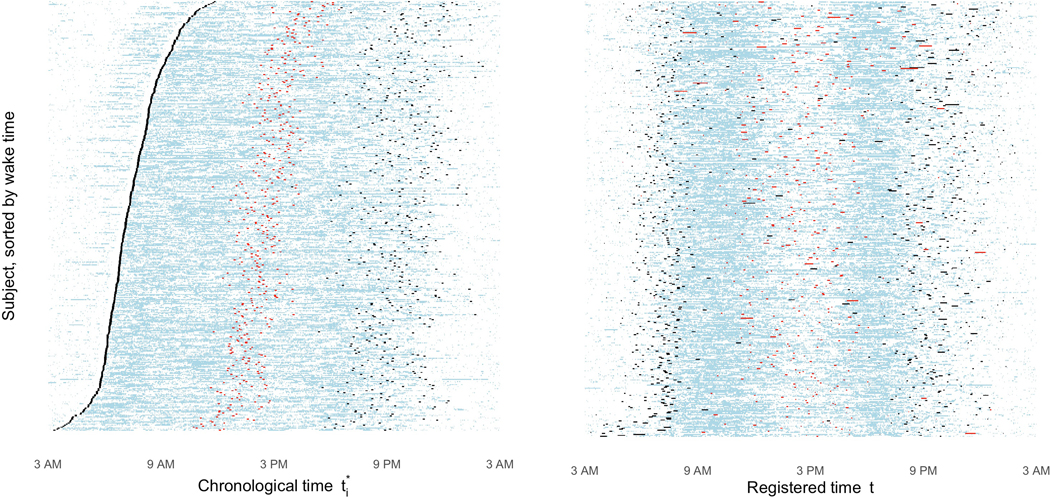
Lasagna plots pre-registration (left) and post-registration (right), with colored markers for landmark times estimated from the (modified) SPT-window approach. In each panel, each row represents a different subject’s binary activity status over time. Blue cells indicate periods of activity, and white cells indicate periods of rest. Sleep and wake times for each subject are marked in black. The daytime interval midpoint (DIM) is marked in red.
3.2.2. Comparing registration with the cosinor model
We fit cosinor curves to each subject’s non-binarized activity count data, noting that the fitted values did not always accurately describe the underlying activity profile, particularly during periods of rest (Supplementary Figure S2). The cosinor models produced three estimated parameters for each subject: the MESOR, the amplitude, and the acrophase. These parameters represent the estimated model-based mean, the peak or maximum deviation from the mean, and the timing of the peak, respectively. Pairwise correlations reveal a strong linear relationship between the MESOR and the amplitude (Pearson correlation coefficient 0.65, Supplementary Figure S1). Examples of potential chronotypes as defined by extreme values of cosinor parameters, analogous to potential chronotypes based on registration parameters presented in Figure 4, are presented in Supplementary Figure S3. Specifically, the cosinor model allows for two main parameter axes on which to evaluate chronotypes. The MESOR and amplitude together describe one’s overall activity intensity, while the acrophase describes horizontal or temporal shifts in one’s activity count profile. Using the cosinor approach, new chronotype discovery is limited because subjects can only deviate along these two axes and their resulting activity count profiles are restricted to the cosine shape.
Below we examine correlations between these cosinor parameters and the two PC scores of registration’s FPCA step. The top row of Figure 7 shows strong negative correlations between the first PC score and the MESOR and amplitude parameters, which is consistent with expectations. All three cosinor parameters showed at least weak associations with at least one slope parameter as well (Supplementary Figure S4); this highlights that, in contrast to the registration parameters, the cosinor parameters are not designed to explicitly explain one source of variability. In the bottom row of Figure 7, no cosinor parameters are correlated with the second PC, suggesting that this is new information gained from registration that the cosinor model could not uncover, perhaps due to its restricting shape.
Figure 7.

Registration-based principal component scores by cosinor parameter values. Registration-based first and second PC scores by cosinor MESOR (top left and bottom left, respectively); registration-based first and second PC scores by cosinor amplitude (top center and bottom center, respectively); and registration-based first and second PC scores by cosinor acrophase (top right and bottom right, respectively).
3.2.3. Comparing registration with unregistered FPCA
For standalone binary FPCA without registration, we explored a range of different numbers of PCs (from 2 to 4) and determined that had the best fit and interpretability. The estimated population-level mean is shown in Figure 8 (black lines). In the left panel, colored lines represent the mean plus (blue) or minus (red) one standard deviation in the first PC. Similarly, the middle and right panels show deviations in the second and third PC’s, respectively. The first PC mostly describes the single-dimensional continuum of morningness vs. eveningness, with some additional information about morning activity intensity and the magnitude of midday dip. The second PC describes overall vertical variability in activity probability. The third PC largely describes the timing of the highest peak, but it also contains some remaining information about wake and sleep times that the first PC failed to capture.
Figure 8.

Population mean activity probability profile from binary FPCA without registration (black lines), plus or minus some variability (red and blue lines, respectively). The left, middle, and right panels demonstrates the mean +/− 1 standard deviation in the first, second, and third principal components, respectively.
Binary FPCA without registration is tasked with making sense of all variability, including both horizontal and vertical. Because of this, both the estimated mean and the types of variability explained in these PCs differ from those of registration (Figure 3), which can capture more advanced levels of variability in patterns of activity probability. Binary FPCA without registration captured the general trend of two activity peaks with a midday dip in its population mean activity probability profile, but these features were not as distinct as those revealed with registration. Furthermore, although its resulting PCs can separate subjects with more active mornings versus more active afternoons (similar to the second PC of registration), this information is confounded by claims about the number of peaks, as well as wake and sleep times.
4. Discussion
We have demonstrated that registration of 24-hour accelerometric rest-activity profiles can provide new insights into the complexity of chronotypes compared to existing methods by carefully separating and handling distinct sources of variability in daily activity patterns. By doing registration, we also uncovered limitations in the interpretability of traditional landmark approaches, specifically the DIM estimated from SPT-windows.
Though registration provides useful information beyond that obtained from the cosinor method, our goal was to introduce registration as an approach that complements rather than replaces the cosinor method. Like the cosinor method, registration reveals interpretable parameters; but registration provides extra flexibility by allowing the shape of the mean activity probability profile to be driven by the data instead of restricted to a cosine-like or any other parametric shape. That said, we acknowledge that some extensions to the cosinor model, including the multiple-component cosinor (Cornelissen, 2014) and the sigmoidally-transformed cosinor (Marler et al., 2006), also provide flexibility that improves upon the original cosinor approach, albeit in different parametric ways. We aim to introduce registration as another available tool among this set of flexible methods, one which learns the shape of the activity probability profile solely from the data.
When comparing registration to the cosinor method in this analysis, it is important to note the difference in outcomes. The cosinor method modeled the log-transformed activity counts as a continuous outcome, whereas the proposed registration method modeled binary states of active vs. rest. We made this modeling choice because we believe that the binarized activity data is of greater utility than the log-transformed (or untransformed) activity counts for studying circadian rhythms. In part, this is because when using the noisy log-transformed counts, modeling methods may be overly influenced by extreme values which may indicate high levels of activity at a particular point in time, but do not convey information about timing of circadian rhythms.
Another limitation of using raw activity counts is the dependence of the results on the definition of activity counts that are often device and manufacturer specific. Binarized accelerometer data has the advantage of being more robust to device-specific activity count definitions. To analyze data from multiple types of devices using our approach, one only needs to establish device-specific thresholds that yield agreeable definitions of active and resting states. We emphasize that this threshold determination is thought of as a pre-processing step rather than part of our proposed registration method. We recommend using established threshold values most appropriate for each device type or manufacturer.
There may still be some circumstances under which continuous activity count data are preferred, for example in younger populations where scientists may want to understand moderate to vigorous physical activity (MVPA). For these research questions, methods for registering Gaussian data have been developed and are available in the “registr” package in R (Wrobel et al., 2020). However, for the reasons listed above, we believe binarized rest-activity profiles are useful for studying circadian rhythms. Our approach to understanding circadian rhythms using binarized accelerometer data is compelling and novel, particularly the insights gained from the second PC obtained with our method.
While registration may seem vastly different from methods commonly used in chronobiology research such as the cosinor method and FPCA, we see many parallels between them. Obviously FPCA and registration are related in the sense that FPCA itself is a step in the registration method. However, a more fundamental similarity across all three methods is the treatment of accelerometer data as functional data across the 24-hour time span. The equations behind the registration, cosinor, and FPCA models are all written as functions of time. From this perspective, we see that the concept of registration is not so far-fetched from the types of methods that are already commonly applied in chronobiology. Finally, our approach is unique in that it borrows information across all subjects. Thus, the approach is potentially more powerful compared to those that focus only on a subject-specific data. This can be leveraged in multiple ways. For example, a population-level algorithm could be developed to more accurately estimate bedtime periods. Additionally, the steepness (fast vs. slow) of the reduction in activity probability in the evening hours after bedtime could be investigated as proxies for sleep inertia.
One limitation of this method, as well as the cosinor method and FPCA, is that they require a full 24 hours of activity data. For older studies these data may not be available since subjects have sometimes been instructed to remove the accelerometer device during sleeping hours (Troiano et al., 2014; Urbanek et al., 2018). However, 24-hour data collection of wrist-worn accelerometer data is becoming increasingly more popular in the physical activity and sleep research communities (Doherty et al., 2017; van Hees et al., 2018). Access to full 24-hour activity profiles will be less of a challenge looking forward.
Relatedly, each of these analysis approaches utilize only a single day of data per subject. We specifically analyzed data collected on a Tuesday for each subject to alleviate day-of-the-week variability, which is minimal in an older population such as the BLSA but may be more relevant for younger populations. Moreover, since our method is only concerned with binarized epochs of activity or rest, our analysis is less sensitive to anomalies such as bouts of exercise on a particular day. That said, to make robust claims about one’s chronotype, one might ideally analyze several days or a week of accelerometer data and estimate within-subject day-to-day stability, variability, and regularity of rest-activity rhythms, topics that are currently under active investigation (Phillips et al., 2017). Work is also underway to create multilevel extensions of the registration method that will address this issue. Although methods targeting multiday registration are not available, the supplementary material contains a sensitivity analysis in which we aggregated over 3 weekdays per subject in an attempt to perform registration on a more stable estimate of one’s weekday activity probability profile. Results of this multiday analysis are similar to those for the single-day analysis, with some changes in the interpretation of PCs. These differences may be due to the reduced sample size, as well as misalignment within subjects across days that is not accounted for in the average curves.
Performing the proposed chronotype analysis on multi-day data, or even single-day data in a larger study population, would require that the proposed method be scalable. Registration is the most computationally demanding element of our approach to identify data-driven chronotypes, and this is indeed scalable: a recent publication applied registration to the UK Biobank dataset (N > 88,000 patients) (Wrobel et al., 2021). Although the analysis is not a study of chronotypes, it involves the registration of daily activity probability profiles from binarized accelerometer data. Several tools were used to ensure that the analysis remained computationally feasible. Although these steps were not necessary for our analysis of the BLSA data (N = 492), they are available in the R package “registr.”
It is also important to note that the interpretations of our registration results, i.e., the potential chronotypes presented in Table 1 and Figure 4, may be specific to this analysis. Performing registration on other datasets or with other analysis specifications, such as the choice of starting point for the 24-hour period, may yield PCs with different interpretations. Here we have defined the daily cycle to start at 3AM; we would expect a different mean function with different PCs and warping functions if the analysis were re-run using a different starting time. Future work is needed to validate our potential new chronotypes using other data sources. As additional future work, it would be interesting to validate the novel measures we introduced in this paper against additional measures of chronotype, such as the MCTQ (Roenneberg et al., 2003, 2007), which were not collected in the BLSA but may be available in other settings.
Our approach assumes that the population has an underlying mean structure that is shared, at least to some extent, across subjects. This shared mean structure is represented in the registration algorithm’s FPCA step through the mean curve and the PCs , which describe the few directions in which subjects generally deviate from that mean curve These are population-level parameters that are shared across subjects, while PC scores model individual differences. In our analysis, the mean curve suggested a morning activity peak, and midday dip, and an afternoon activity peak, with rest during nighttime hours. This biphasic structure has also been identified as an FPCA component of other large-scale accelerometer studies such as the National Health and Nutrition Examination Survey (NHANES) (Leroux et al., 2019) and the Osteoporotic Fractures in Men (MrOS) study (Zeitzer et al., 2018) as well as in monkeys (Leroux, 2015). Furthermore, the existence of a shared mean structure is supported by the BLSA data in the sense that many warping functions lie close to the identity line, suggesting minimal warping required to align these subjects. A shared mean structure may not be relevant to individuals with highly irregular chronotypes, such as shift workers or patients with sleep disorders, and it may be possible to identify these individuals using either warping functions or subject-level goodness-of-fit analyses. In general, registration is not appropriate for settings in which behavioral patterns are very dissimilar across participants. In these cases, binary FPCA cannot identify useful PCs with which to construct subject-specific mean templates – because shared patterns don’t exist across subjects.
Our method may be an important tool for identifying more subtle deviations from the mean profile of a typical population. Chronotype differences have been shown to relate to health outcomes even among more normally-phased healthy participants. Approximately 87% of day workers suffer from social jetlag, which is akin to a milder form of shift work. Furthermore, about a quarter of the general population meet the criteria for advanced sleep phase disorder or delayed sleep phase disorder, defined as deviations of at least 2 hours from desired sleep time (Roenneberg & Merrow, 2016). Subtle differences in circadian rhythms such as social jetlag or <7 hours of sleep per night have been linked to health outcomes including obesity, addiction, and cognitive under-performance (Goel et al., 2015; Wittmann et al., 2006). This suggests the clinical utility of our registration method, which we have shown can identify more nuanced chronotypes in a somewhat homogeneous population than other approaches. The registration-based chronotypes can be defined on a four-dimensional continuum, similar to but more flexible than the well-known single-dimensional morningness-eveningness chronotype continuum, which has been shown to correlate with health outcomes (Partonen, 2015). Future work is needed to both validate these registration-based dimensions using independent data sources and to explore associations between these potential chronotypes and health outcomes.
Supplementary Material
Acknowledgments
Funding
This work was supported in part by the National Institutes of Health under Grant R01NS097423.
J.A.S. is supported by the National Institutes of Health under Grant R01AG061786 and Grant U01AG057545.
V.Z. is supported by the National Institutes of Health under Grant R01AG054771 and Grant U01AG057545.
Data from this study were obtained from the Baltimore Longitudinal Study of Aging, a study of the Intramural Research Program of the National Institute on Aging.
Abbreviations
- BLSA
Baltimore Longitudinal Study of Aging
- DIM
Daytime interval midpoint
- FPCA
Functional principal components analysis
- HRR
Heart rate reserve
- MCTQ
Munich ChronoType Questionaire
- MEQ
Morningness-Eveningness Questionnaire
- MESOR
Midline estimating statistic of rhythm
- MrOS
Osteoporotic Fractures in Men study
- MVPA
Moderate to vigorous physical activity
- NHANES
National Health and Nutrition Examination Survey
- PC
Principal component
- SPT-window
Sleep period time window
Footnotes
Data availability statement
Data from the BLSA are available upon request from the BLSA website [https://www.blsa.nih.gov/]. All requests are reviewed by the BLSA Data Sharing Proposal Review Committee.
Disclosure of interest
The authors report no conflict of interest.
References
- Cornelissen G. (2014). Cosinor-based rhythmometry. Theoretical Biology and Medical Modelling, 11(16), 1–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doherty A, Jackson D, Hammerla N, Plötz T, Olivier P, Granat MH, White T, Van Hees VT, Trenell MI, Owen CG, Preece SJ, Gillions R, Sheard S, Peakman T, Brage S, & Wareham NJ (2017). Large scale population assessment of physical activity using wrist worn accelerometers: The UK biobank study. PLoS One, 12(2), 1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunlap JC, Loros JJ, & DeCoursey PJ (2004). Chronobiology: Biological timekeeping. Sinauer Associates. [Google Scholar]
- Gershon A, Kaufmann CN, Depp CA, Miller S, Do D, Zeitzer JM, & Ketter TA (2018). Subjective versus objective evening chronotypes in bipolar disorder. Journal of Affective Disorders, 225, 342–349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gershon A, Ram N, Johnson SL, Harvey AG, & Zeitzer JM (2016). Daily actigraphy profiles distinguish depressive and interepisode states in bipolar disorder. Clinical Psychological Science, 4(4), 641–650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goel N, Basner M, & Dinges DF (2015). Phenotyping of neurobehavioral vulnerability to circadian phase during sleep loss. Methods in enzymology 552, 285–308. [DOI] [PubMed] [Google Scholar]
- Goldsmith J, Zipunnikov V, & Schrack J. (2015). Generalized multilevel function-on-scalar regression and principal component analysis. Biometrics, 71(2), 344–353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halberg F, Tong YL, & Johnson EA (1967). Circadian system phase - An aspect of temporal morphology; procedures and illustrative examples. In von Mayersbach H. (Ed.), The Cellular Aspects of Biorhythms (pp. 20–48). Springer-Verlag. [Google Scholar]
- Horne JA, & Ostberg O. (1976). A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. International Journal of Chronobiology, 4(2), 97–110. [PubMed] [Google Scholar]
- Jones SE, van Hees VT, Mazzotti DR, Marques-Vidal P, Sabia S, van der Spek A, Dashti HS, Engmann J, Kocevska D, Tyrrell J, Beaumont RN, Hillsdon M, Ruth KS, Tuke MA, Yaghootkar H, Sharp SA, Ji Y, Harrison JW, Freathy RM, … Wood AR (2019). Genetic studies of accelerometer-based sleep measures yield new insights into human sleep behaviour. Nature Communications, 10(1), 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufmann CN, Gershon A, Depp CA, Miller S, Zeitzer JM, & Ketter TA (2018). Daytime midpoint as a digital biomarker for chronotype in bipolar disorder. Journal of Affective Disorders, 241, 586–591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leroux A. (2015). Activity patterns in a Parkinson’s monkey model (Issue April) [Masters Thesis, Johns Hopkins University]. https://jscholarship.library.jhu.edu/handle/1774.2/60374 [Google Scholar]
- Leroux A, Di J, Smirnova E, Mcguffey EJ, Cao Q, Bayatmokhtari E, Tabacu L, Zipunnikov V, Urbanek JK, & Crainiceanu C. (2019). Organizing and analyzing the activity data in NHANES. Statistics in Biosciences, 11(2), 262–287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marler MR, Gehrman P, Martin JL, & Ancoli-Israel S. (2006). The sigmoidally transformed cosine curve: A mathematical model for circadian rhythms with symmetric non-sinusoidal shapes. Statistics in Medicine, 25(22), 3893–3904. [DOI] [PubMed] [Google Scholar]
- Melo MCA, Abreu RLC, Linhares Neto VB, de Bruin PFC, & de Bruin VMS (2017). Chronotype and circadian rhythm in bipolar disorder: A systematic review. Sleep Medicine Reviews, 34, 46–58. [DOI] [PubMed] [Google Scholar]
- Partonen T. (2015). Chronotype and health outcomes. Current Sleep Medicine Reports, 1(4), 205–211. [Google Scholar]
- Phillips AJK, Clerx WM, O’Brien CS, Sano A, Barger LK, Picard RW, Lockley SW, Klerman EB, & Czeisler CA (2017). Irregular sleep / wake patterns are associated with poorer academic performance and delayed circadian and sleep / wake timing. Scientific Reports, 7(1), 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramsay JO, & Silverman BW (2002). Applied Functional Data Analysis: Methods and Case Studies (1st ed.). Springer-Verlag. 10.1111/j.1467-985x.2004.t01-5-.x [DOI] [Google Scholar]
- Ramsay JO, & Silverman BW (2005). Functional data analysis (2nd ed.). Springer-Verlag. [Google Scholar]
- Refinetti R, Cornélissen G, & Halberg F. (2007). Procedures for numerical analysis of circadian rhythms. Biological Rhythm Research, 38(4), 275–325 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roenneberg T, Kuehnle T, Juda M, Kantermann T, Allebrandt K, Gordijn M, & Merrow M. (2007). Epidemiology of the human circadian clock. Sleep Medicine Reviews, 11(6), 429–438. [DOI] [PubMed] [Google Scholar]
- Roenneberg T, & Merrow M. (2016). The circadian clock and human health. Current Biology, 26(10), R432–R443. 10.1016/j.cub.2016.04.011 [DOI] [PubMed] [Google Scholar]
- Roenneberg T, Wirz-Justice A, & Merrow M. (2003). Life between clocks: Daily temporal patterns of human chronotypes. Journal of Biological Rhythms, 18(1), 80–90. [DOI] [PubMed] [Google Scholar]
- Roveda E, Vitale J, Montaruli A, Galasso L, Carandente F, & Caumo A. (2017). Predicting the actigraphy-based acrophase using the Morningness–Eveningness Questionnaire (MEQ) in college students of North Italy. Chronobiology International, 34(5), 551–562. [DOI] [PubMed] [Google Scholar]
- Schrack JA, Kuo PL, Wanigatunga AA, Di J, Simonsick EM, Spira AP, Ferrucci L, & Zipunnikov V. (2019). Active-to-sedentary behavior transitions, fatigability, and physical functioning in older adults. The Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 74(4), 560–567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schrack JA, Leroux A, Fleg JL, Zipunnikov V, Simonsick EM, Studenski SA, Crainiceanu C, & Ferrucci L. (2018). Using heart rate and accelerometry to define quantity and intensity of physical activity in older adults. The Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 73(5), 668–675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schrack JA, Zipunnikov V, Goldsmith J, Bai J, Simonsick EM, Crainiceanu C, & Ferrucci L. (2014). Assessing the physical cliff: Detailed quantification of age-related differences in daily patterns of physical activity. The Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 69(8), 973–979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swihart BJ, Caffo B, James BD, Strand M, Schwartz BS, & Punjabi NM (2010). Lasagna plots : A saucy alternative to spaghetti plots. Epidemiology, 21(5), 621–625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Troiano RP, McClain JJ, Brychta RJ, & Chen KY (2014). Evolution of accelerometer methods for physical activity research. British Journal of Sports Medicine, 48(13), 1019–1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urbanek JK, Spira AP, Di J, Leroux A, Crainiceanu C, & Zipunnikov V. (2018). Epidemiology of objectively measured bedtime and chronotype in US adolescents and adults: NHANES 2003–2006. Chronobiology International, 35(3), 416–434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Linde A. (2008). Variational Bayesian functional PCA. Computational Statistics and Data Analysis, 53(2), 517–533. [Google Scholar]
- van Hees VT, Sabia S, Anderson KN, Denton SJ, Oliver J, Catt M, Abell JG, Kivimäki M, Trenell MI, & Singh-Manoux A. (2015). A novel, open access method to assess sleep duration using a wrist-worn accelerometer. PLoS One, 10(11), 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Hees VT, Sabia S, Jones SE, Wood AR, Anderson KN, Kivimäki M, Frayling TM, Pack AI, Bucan M, Trenell MI, Mazzotti DR, Gehrman PR, Singh-Manoux BA, & Weedon MN (2018). Estimating sleep parameters using an accelerometer without sleep diary. Scientific Reports, 8(1), 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wanigatunga AA, Di J, Zipunnikov V, Urbanek JK, Kuo P-L, Simonsick EM, Ferrucci L, & Schrack JA (2019). Association of total daily physical activity and fragmented physical activity with mortality in older adults. JAMA Network Open, 2(10), e1912352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witting W, Kwa IH, Eikelenboom P, Mirmiran M, & Swaab DF (1990). Alterations in the circadian rest-activity rhythm in aging and Alzheimer’s disease. Biological Psychiatry, 27(6), 563–572. [DOI] [PubMed] [Google Scholar]
- Wittmann M, Dinich J, Merrow M, & Roenneberg T. (2006). Social jetlag: Misalignment of biological and social time. Chronobiology International, 23(1–2), 497–509. [DOI] [PubMed] [Google Scholar]
- Wrobel J, Goldsmith J, & McDonnell E. (2020). registr: Curve registration for exponential family functional data (R package version 1.0.0). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wrobel J, Muschelli J, & Leroux A. (2021). Diurnal physical activity patterns across ages in a large uk based cohort: The uk biobank study. Sensors, 21(4), 1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wrobel J, Zipunnikov V, Schrack J, & Goldsmith J. (2019). Registration for exponential family functional data. Biometrics, 75(1), 48–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zee PC, Attarian H, & Videnovic A. (2013). Circadian rhythm abnormalities. Continuum (Minneapolis, Minn), 19(1), 132–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeitzer JM, Blackwell T, Hoffman AR, Cummings S, Ancoli-Israel S, & Stone K. (2018). Daily patterns of accelerometer activity predict changes in sleep, cognition, and mortality in older men. The Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 73(5), 682–687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeitzer JM, David R, Friedman L, Mulin E, Garcia R, Wang J, Yesavage JA, Robert PH, & Shannon W. (2013). Phenotyping apathy in individuals with Alzheimer disease using functional principal component analysis. The American Journal of Geriatric Psychiatry, 21(4), 391–397. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.