

Abstract
Global climate change has led to range shifts in plants and animals, thus threatening biodiversity. Latitudinal shifts have been shown to be more pronounced than elevational shifts, implying that northern range edge margins may be more capable to keeping pace with warming than upper elevational limits. Additionally, global climate change is expected to disadvantage habitat specialists. In the Alps, climatic variation along the elevation gradient allows the coexistence of habitat specialists and generalists. Alpine species are anticipated to adapt their elevational ranges to the change of various climate variables caused by global climate change. Regional differences might buffer elevational shifts. Furthermore, distinct climate variables might differently affect the shifts of habitat specialists and generalists. To study the effect of climate change on Alpine species, we analysed hunting bag, climate and biogeographical data of two grouse species (Tetrao tetrix and Lagopus muta) and two hare species (Lepus timidus varronis and L. europaeus) in Grisons, Switzerland, over a period of 30 years. Our results based on 84,630 harvested specimens were as follows: (1) only three out of seven climate variables changed significantly within the study period. (2) The grouse species significantly shifted towards higher elevations, whereas the hare species only shifted in their minimum/maximum elevations. (3) Hunting elevation of habitat generalists increased more than in habitat specialists. (4) The elevational shifts were mostly related to the number of frost days. (5) Hunting elevation increased especially in the southern biogeographical region. To conclude, all four taxa respond to climate change but habitat generalists more rapidly than habitat specialists. The range shift to higher elevations due to global climate change will lead to a reduction in habitat availability for snow‐adapted species. Climate change is thus a serious threat to alpine biodiversity. Regions rich in alpine habitats will have an increased responsibility to conserve these species.
Keywords: Alpine mountain hare, black grouse, climate warming, European hare, interspecific competition, rock ptarmigan
Alpine species are anticipated to adapt their elevational ranges to the change of various climate variables caused by global climate change. We analysed hunting bag, climate and biogeographical data of two grouse species (Tetrao tetrix and Lagopus muta) and two hare species (Lepus timidus varronis and L. europaeus) in Grisons, Switzerland, over a period of 30 years. All four taxa respond to climate change but habitat generalists more rapidly than habitat specialists. The range shift to higher elevations due to global climate change will lead to a reduction in habitat availability for snow‐adapted species.

1. INTRODUCTION
The impacts of global climate change on the future of biodiversity are likely tremendous (Bellard et al., 2012). Species respond to climate change through phenotypic plasticity or evolutionary responses. For instance, during the recent rapid climate change, plant and animal species shifted their geographical ranges both latitudinally and altitudinally (Chen et al., 2011; Gottfried et al., 2012; Harsch et al., 2009; Johnson et al., 2010; Lenoir et al., 2008; Lu et al., 2021; Parmesan & Yohe, 2003; Pauli et al., 2012; Post et al., 2009; Thomas et al., 2004). On average, species moved away from the Equator at a median rate of 6.0–16.9 km decade−1. In parallel, species shifted towards higher elevations at a rate of 6.1–11.0 m decade−1 (Chen et al., 2011). Despite the statistical link between range shifts and warming (Chen et al., 2011), and although distances needed to track climate effects are much shorter for altitudinal than for latitudinal shifts (Loarie et al., 2009), there is a lag in demonstrated elevational responses to increasing temperatures. This implies that species in montane habitats respond less to climate change than other species, and may therefore be more vulnerable to extinction. The lag in elevational response might be because suitable conditions cannot be reached easily in montane habitats (as peaks are isolated), or because mountainous terrain is characterized by topographic and microclimatic complexity which masks the effect of climate change (Chen et al., 2011). In line with this, it has been suggested that the apparent lag in previous research may be due to unbalanced representation of taxa and their habitats (Reif & Flousek, 2012). For example, taxa included in meta‐analyses (Chen et al., 2011; Parmesan & Yohe, 2003) were primarily plants and arthropods with limited ability to respond to climate change due to vegetative reproduction (e.g. grasses and herbs), long generation times (e.g. trees) or narrow ecological niches (e.g. tree‐dependent insects). In mountain ungulate species characterized by a high mobility, pronounced effects of climate change, that is, range shifts towards higher elevations, have been demonstrated (Büntgen et al., 2017). But how are less mobile medium‐sized species, which inhabit open or semi‐open montane and alpine areas, and are not dependent on nest holes or burrows but can choose their home range according to the availability of food and shelter, responding to climate warming? We expect to find noticeable effects of climate change in medium‐sized mammal and bird species in terms of range shifts towards higher elevations as well.
Among medium‐sized species living in the Alps, there are species that are adapted to cold conditions and, thus, specialized in alpine habitats, whereas others are more generalist regarding their habitat requirements. One adaptation of specialists to cold and snowy conditions is a colour change to white during winter which exhibit both the Alpine mountain hare (Lepus timidus varronis) and the rock ptarmigan (Lagopus muta), whereas generalists such as the European hare (L. europaeus) and the black grouse (Tetrao tetrix) do not show this adaptive trait. European hares evolved in steppe habitats and now occur in diverse open or semi‐open habitats, from arable land to salt marshes (Hackländer & Schai‐Braun, 2018). The black grouse is found in open or semi‐open landscapes both in lowland areas and above the tree line (Balzari & Gygax, 2019). With increasing temperatures, habitats at higher elevations will become increasingly available to them although the available surface area of specific habitats, that is, timberline ecotone will be decreasing as well. The European hare has already been shown to inhabit high elevations in the Alps (Schenker et al., 2020), also during winter. Hence, generalists seem to expand their range to include higher elevation sites due to climate warming in the Alps. Whether the expected elevational shifts to higher elevations in the Alps are differing in their velocity between specialists and generalists is yet unknown but might be decisive for the outcome of competition between congeneric species such as the two lagomorph species with high potential for niche overlap (Gidoin et al., 2015).
Recent patterns of anthropogenic global climate change and their underlying drivers are well established (IPCC, 2014, 2021). However, it is unclear which of numerous climate variables might have an effect on Alpine species’ elevational shifts. In the first step, climate variables have to be identified that are changing markedly in the habitat and within the time period studied in order to be potentially responsible for the elevational shifts of Alpine species. In the second step, the impact of the changing climate variables on the Alpine species’ elevational shifts has to be investigated.
Local climate may mitigate the impact of climate warming (Feldmeier et al., 2020). Especially, a heterogeneous mountain topography may provide microclimatic conditions that can buffer macroclimatic warming as has been shown for the Alpine salamander (Salamandra atra) and the common lizard (Zootoca vivipara; Feldmeier et al., 2020). The canton of Grisons is biogeographically particularly rich encompassing three of the six existing biogeographical regions in Switzerland (Gonseth et al., 2001). As each locality has been biogeographically classified based on patterns of distribution of flora and fauna, Alpine species might respond to climate warming differently based on the biogeographical regions they are inhabiting. Elevational shifts might therefore vary due to the biogeographical regions in Grisons.
The goal of this study was to investigate the elevational shift of four species (two habitat specialists and two generalists, two bird species and two mammal species) living in the Alps over three decades. Our hypotheses were as follows: (1) several climate variables measured during the last three decades (our study period) demonstrate climate warming in Grisons; (2) elevational ranges of two hare and grouse species increased within the last three decades in the Alps; (3) elevational shifts are differing in their velocity in habitat specialists and generalists inhabiting the Alps; (4) elevational range shifts within the last three decades in two hare and grouse species in the Alps are affected by particular climate variables as well as (5) biogeographical regions in Grisons. To test these hypotheses, we analysed hunting bag, climate and biogeographical data of four species living in the Alps in Grisons over a period of 30 years.
2. METHODS
2.1. Data collection
To test our hypothesis, we focus on four species which fulfil these criteria and occur in the Swiss canton Grisons, a central Alpine region with elevations from 260 to 4049 m a.s.l.: two lagomorphs, the mountain hare and the European hare as well as two tetraonids, the rock ptarmigan and the black grouse. Here, mountain hares inhabit open and semi‐open alpine habitats mainly above the tree line, that is, from 800 to 3100 m a.s.l.; European hares inhabit alpine valleys at lower elevations up to 2800 m a.s.l. (Averianov et al., 2003). Rock ptarmigan inhabit rocky, treeless alpine regions between 1900 and 2600 m a.s.l.; black grouse inhabit semi‐open habitats and richly structured habitats with open spaces, preferably in a small range along the timberline between 1500 and 2200 m a.s.l. (Balzari & Gygax, 2019). Climate change has been pronounced in Grisons: since 1961 temperatures have increased significantly in winter (0.3–0.4°C decade−1) as well as in summer (0.4–0.5 decade−1), and the annual number of frost days has decreased significantly by up to 10 decade−1. The 0°C isotherm has moved upwards at a rate of 30–70 m decade−1, that is, shifted approximately 150–200 m per 1°C (MeteoSchweiz, 2012).
Since their introduction in the year 1991, hunting regulations are used as a major tool for wildlife management decisions in Grisons. Important areas were placed under hunting protection and personal quotas per hunter were reduced. For all four studied species, monitoring and control instruments are available to monitor both the population status and exploitation by hunting. Game sanctuaries (areas with a hunting ban) for all four species exist on more than 900 km2. In winter, game refuges of an equivalent area exist in order to prevent disturbance of these species, especially by winter tourism. Information on the elevational ranges of the four species in fall were collected from licensed hunters. In Grisons, hares are harvested from 1 October to 20 November each year and tetraonids from 16 October to 30 November. Licence holders are allowed to hunt anywhere within the canton except in the game sanctuaries; motor vehicle use is restricted. Hence, hunters start climbing up the hill from the place where the use of a motor vehicle is prohibited (village, pass summit) and hunt the game they have a permit for. Each harvested animal must be registered in a shooting list immediately after shooting, together with all data on date, time, place and elevation. Overharvesting is also prevented by issuing a quota based on population estimates prior to the hunting period.
We assumed that if our hypotheses were correct, hunters would have to follow their shifting quarry towards higher elevations so that sites at which animals were harvested would shift in elevation over time. To test the hypotheses, we analysed the elevation (m a.s.l.) at which hunters harvested game from a dataset of obligate hunters reports on their bag during a period of 30 years (1991–2020). Using hunting bag data as proxy for hare abundance has its limitations as the relationship is rarely straightforward (Langbein et al., 1999). However, the utility of long‐term time‐series of (hare) hunting game bags for ecological and climate change studies has been shown already (Reid et al., 2021). Elevations were determined either from contours on maps (1:25,000) or with the help of a GPS receiver or a hunting app (JagdGR, www.jagdgr.ch, dynamicstudio GmbH, Poschiavo) to the nearest 50 m depending on the hunter's equipment; information was verified by official game wardens. Because the elevation is an important steering tool for regulating hunting pressure in the hunting regulations in Grisons since 1991, the hunters are very familiar with the elevation conditions in their hunting district. Mean annual ambient temperature (5 cm above grass, °C), annual snow depth (cm), annual fresh snow depth (cm), annual number of frost days (minimum temperature <0°C) and annual number of days with snow reliability (snow depth ≥30 cm) for the canton of Grisons were calculated by averaging data from four weather stations in Grisons above 1200 m a.s.l. (Davos 46°49′N, 9°51′E, Samedan 46°32′N, 9°53′E, Scuol 46°48′N, 10°17′E, San Bernardino 46°28′N, 9°11′E; data provided by meteoswiss, for the data used to calculate the climate variables see Supplemental Materials S1, S2 and S3). Furthermore, mean ambient temperature (°C) and number of frost days for the months October and November (hunting period) were determined. The canton of Grisons encompasses three of the six existing biogeographical regions in Switzerland (Gonseth et al., 2001). The biogeographical classification of Switzerland employs a purely statistical approach, based on patterns of distribution of flora and fauna, and adapted to communal boundaries (Gonseth et al., 2001). As within a relatively small region such contrasting regions that differ in flora and fauna can be found, the canton of Grisons provides a good case study to investigate the responses of alpine species to global climate change. The Office for Hunting and Fishing of Grisons (Amt für Jagd und Fischerei Graubünden) has elaborated a slightly different assignment of the same three biogeographical regions for each hunting district (a smaller resolution as the communal borders) based on the specific weather conditions created by the structure of the mountains and the dominant exposition of the hunting district to the Atlantic, insubric and continental weather conditions (Just, 2013; for a map of the classification of the biogeographical regions in Grisons by the Office for Hunting and Fishing of Grisons, see Supplemental Material S4, for the hunting bag data including hunting districts and biogeographical regions, see Supplemental Material S5), which we use for our data analysis. Note that the different hunting districts do not differ in hunting management.
2.2. Data analysis
All analyses were computed with the software R 4.1.0 (R Development Core Team, 2019). We analysed the hunting bag and climate data by fitting linear regression models. All models analysing time series of original data (several thousand per year) included an autoregressive (AR1) error structure to explicitly account for temporal autocorrelation. All corrections for autocorrelation were successful, as checked with the R package ‘itsadug’ (van Rij et al., 2020). To compute linear regressions, we used the linear, parametric part of bam models (R package mgcv, Wood, 2004), because bam allows for AR1 error structures within years (while function lme::gls ran out of memory). When spline functions are not used, as was the case here, function bam returns coefficients, t‐ and p‐values that are identical to other ordinary least squares functions, such as lm.
To examine the effect of climate variables and biogeographical region on the hunting elevation, we detrended the time series by differencing subsequent years. As some climatic variables were only available as means per year, we used yearly averages of hunting elevations, and detrending was based on these means. Yearly means were not significantly autocorrelated. Using this approach, we asked whether extreme shifts in hunting elevation were caused by extreme changes in climate, compared with the previous year. Note that in this case, the estimation of the hunting elevation was still based on a sample size of several thousand per year.
First, climate change in the study area was measured by examining every climate variable as response variable for the predictor variable year (corrected for autocorrelation). Second, the response variable hunting elevation (original values) was examined by fitting the predictor variable year for each species separately, again correcting for autocorrelation. Additionally, the response variables minimum (≤15th percentile) and maximum (≥85th percentile) hunting elevation were investigated for the predictor variable year. Third, the effects of climate on the annual means of the hunting elevations were examined by fitting a linear regression model on detrended data for each species separately. The full models for the response variables hunting elevation included the different climate variables as predictor variables and their two‐way interaction terms. Multicollinearity between the predictor variables was checked by calculating the variance inflation factor (VIF) with R package ‘car’ (Fox & Weisberg, 2019) for all predictor variables in each model. There was no VIF > 5.2. The full models were used to choose the models that described best the data based on delta AIC using the R package ‘MuMln’ (Bartoń, 2013).
Following Zuur et al. (2009), we subsequently determined whether predictors significantly contributed to the variance explained by the best model. p‐values and estimates (β) were extracted from the best model. Fourth, the response variable hunting elevation (original values) was examined by fitting a linear regression model for the predictor variable biogeographical region (factor with levels northern, central and southern) for each species separately to examine the effect of the biogeographical region in Grisons on the hunting elevation. Post‐hoc tests were computed for the biogeographical region using the Tukey's all‐pair comparisons method in the R package ‘multcomp’ (Hothorn et al., 2008). Fifth, the response variable hunting elevation (original values, corrected for autocorrelation) was examined as a function for the predictor variable year and biogeographical region for each species. Sixth, the effects of climate on the annual means of the hunting elevations in the different biogeographical regions were examined by fitting a linear regression model on detrended data for each species separately. The full models for the response variables hunting elevation included the two‐way interaction between the different climate variables and the biogeographical region as predictor variables. Multicollinearity between the predictor variables was again checked calculating VIF for all predictor variables in each model. There was no VIF > 5.2. p‐values and estimates (β) were extracted from the full model. We visually checked the normality of the model residuals by means of a normal probability plot and histograms. Homogeneity of variances and goodness‐of‐fit were examined by plotting residuals versus fitted values (Faraway, 2006).
3. RESULTS
3.1. Climate warming in the canton of Grisons
Mean annual ambient temperature measured at all four weather stations increased significantly (Davos: +0.3°C decade−1, R 2 = .27, p < .001, n = 29; Samedan: +0.8°C decade−1, R 2 = .66, p < .001, n = 30; San Bernardino: +0.5°C decade−1, R 2 = .39, p < .001, n = 29; Scuol: +0.5°C decade−1, R 2 = .35, p < .001, n = 28), whereas annual number of frost days decreased significantly during the study period except for the weather station Scuol (Davos: −9.2 days decade−1, R 2 = .12, p = .017, n = 30; Samedan: −15.2 days decade−1, R 2 = .59, p < .001, n = 30; San Bernardino: −13.2 days decade−1, R 2 = .26, p = .008, n = 30; Scuol: p > .10; Figure 1). Mean ambient temperature in October and November measured at all four weather stations increased significantly as well (Davos: +0.6°C decade−1, R 2 = .19, p = .010, n = 29; Samedan: +0.8°C decade−1, R 2 = .34, p < .001, n = 30; San Bernardino: +0.6°C decade−1, R 2 = .21, p = .005, n = 29; Scuol: +0.5°C decade−1, R 2 = .11, p = .036, n = 29). The number of frost days in October and November, snow depth, fresh snow depth and annual number of days with snow reliability did not significantly correlate with years during the study period (p > .10).
FIGURE 1.
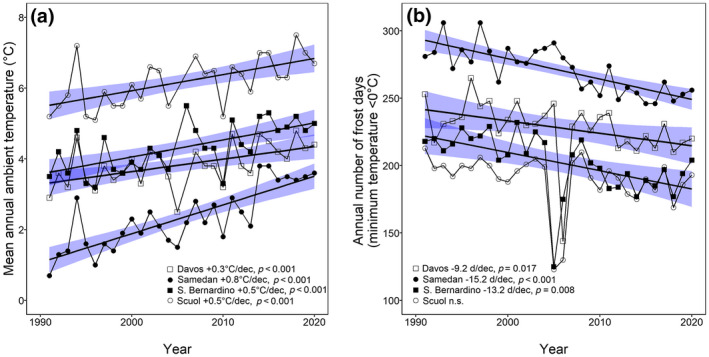
(a) Mean annual ambient temperature (°C) and (b) annual number of frost days (minimum temperature <0°C) for four weather stations in Grisons above 1200 m a.s.l. during the 30 study years (1991–2020). The lines represent the regression lines (statistically significant) with the blue area displaying the confidence intervals. Model corrected for autocorrelated errors
3.2. Elevational shift in harvesting elevation
In total, we obtained the elevation at which 84,630 specimens were harvested (black grouse: n = 4551; rock ptarmigan: n = 11,963; European hare: n = 39,285; Alpine mountain hare: n = 28,831). For the two grouse species, we found a significant shift towards higher elevations within the study period, whereas no significant shift was noticeable in the European hare (p > .05) and the Alpine mountain hare (p > .10; Figure 2). The shift in black grouse harvesting sites of 53 m decade−1 was more pronounced (R 2 = .05, p < .001) than in the rock ptarmigan with a shift of 33 m decade−1 (R 2 = .02, p < .001). The minimum hunting elevation of all four species increased significantly (rock ptarmigan: 57 m −1, R 2 = .11, p < .001, n = 2217; black grouse: 61 m decade−1, R 2 = .13, p < .001, n = 862; Alpine mountain hare: 28 m decade−1, R 2 = .02, p < .001, n = 5070; European hare: 31 m decade−1, R 2 = .02, p < .001, n = 6429). The correlation between year and maximum hunting elevation differed between species, especially in the two hare species (rock ptarmigan: 17 m −1, R 2 = .03, p = .015, n = 2138; black grouse: 36 m decade−1, R 2 = .10, p < .001, n = 779; Alpine mountain hare: not significant, p > .10, n = 4546; European hare: −16 m decade−1, R 2 = −.005, p = .037, n = 6611).
FIGURE 2.
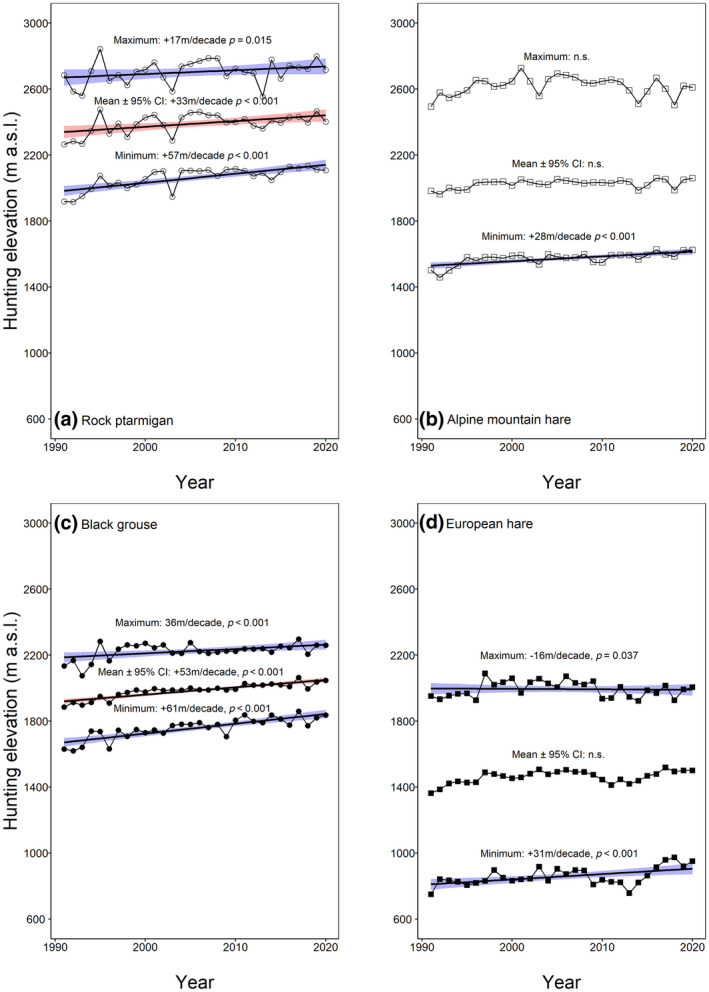
Harvesting elevation (m a.s.l.), minimum (≤15th percentile, m a.s.l.) and maximum hunting elevation (≥85th percentile, m a.s.l.) of the (a) rock ptarmigan (n = 11,963, n min = 2217, n max = 2138), (b) Alpine mountain hare (n = 28,831, n min = 5070, n max = 4546), (c) black grouse (n = 4551, n min = 862, n max = 779) and (d) European hare (n = 39,285, n min = 6429, n max =6611) collected over a 30‐year period (1991–2020) during the yearly hunting period in autumn in Grisons. The symbols represent the mean. The lines represent the regression lines (statistically significant) with the red/blue area displaying the confidence intervals. Modelled on original data, corrected for autocorrelated errors
3.3. Effect of climate warming on elevational shifts of harvesting sites
Detrended climate variables had a significant effect in both grouse species and in the Alpine mountain hare (Figure 3; for an overview of all model coefficients, see Supplemental Material S6). The hunting elevation of the rock ptarmigan was positively affected by the annual fresh snow depth (β = 0.386, R 2 = .37, p = .005; Figure 3a) but negatively affected by the number of frost days in October and November (β = −9.013, R 2 = .37, p = .006; Figure 3b). The hunting elevation of the Alpine mountain hare was negatively influenced by the mean annual ambient temperature (β = −14.513, R 2 = .40, p = .007; Figure 3c) and positively influenced by the number of frost days in October and November (β = 3.669, R 2 = .40, p = .004; Figure 3d). The hunting elevation of the black grouse was positively influenced by the annual fresh snow depth (β = 0.159, R 2 = .35, p = .006, Figure 3e) and the interaction of annual number of frost days and annual snow depth (β = 0.137, R 2 = .35, p = .003; Figure 3f). No climate variable was significantly affecting the hunting elevation of the European hare (number of frost days in October and November: p > .05, Figure 3g, all other climate variables: p > .10).
FIGURE 3.

Relationships between different climate variables and the hunting elevation (m a.s.l.) of the four species collected over a 30‐year period (1991–2020) during the yearly hunting period in autumn in Grisons. Both hunting elevation and climate variables were detrended by yearly differencing. Effect of differences in (a) annual fresh snow depth (cm) and (b) number of frost days in October and November on relative elevation of rock ptarmigan, (c) mean annual ambient temperature (°C), (d) number of frost days in October and November on elevation in Alpine mountain hare, (e) annual fresh snow depth (cm), (f) the interaction of annual number of frost days and annual snow depth (cm) on elevation in black grouse and (g) number of frost days in October and November on elevation in European hare. The lines represent the regression lines (statistically significant) with the red/blue area showing the confidence intervals. Modelled on annual means of the data
3.4. Effect of biogeographical region on elevational shifts of harvesting sites
The biogeographical region influenced significantly the harvesting elevation of all four species (for an overview of post‐hoc test results, see Supplemental Material S7). Harvesting elevation increased in all four species from the northern to the central to the southern region in Grisons (Figure 4). Hunting elevation of the Alpine mountain hare increased significantly in the northern and southern biogeographical region (Figure 5a–c, for an overview of the model coefficient [parameter estimates β and p‐values], see Supplemental Material S8), whereas no significant annual effect in the European hare was noticeable (p > .10). Hunting elevation in the two grouse species increased significantly in all biogeographical regions of Grisons (Figure 5d–f) except in the rock ptarmigan in the northern biogeographical region. In both bird species, the increase was highest in the southern biogeographical region.
FIGURE 4.

Hunting elevation (m a.s.l., medians with 25th/75th and 10th/90th percentiles) of the two (a) hare species and (b) grouse species in northern, central and southern biogeographical regions in Grisons collected over a 30‐year period (1991–2020). White: Alpine mountain hare (n = 28,831) and rock ptarmigan (n = 11,963), grey: European hare (n = 39,285) and black grouse (n = 4551). Different letters indicate significant differences among groups (post‐hoc: p < .001). Modelled on original data
FIGURE 5.
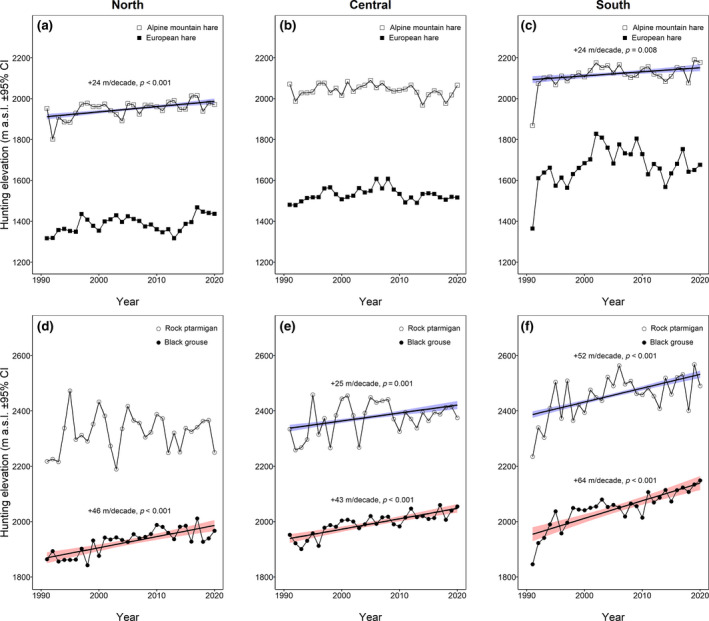
Hunting elevation (m a.s.l.) at which the two (a, b, c) hare species and (d, e, f) grouse species were harvested in (a, d) northern, (b, e) central and (c, f) southern biogeographical regions in Grisons collected over a 30‐year period (1991–2020) during the yearly hunting period in autumn. Open rectangles: Alpine mountain hare (n = 28,831), filled rectangles: European hare (n = 39,285), open circles: rock ptarmigan (n = 11,963), filled circles: black grouse (n = 4551). The symbols represent the mean. The lines represent the regression lines (statistically significant) with the blue/red area showing the confidence intervals. Modelled on original data, corrected for autocorrelated errors
3.5. Regional differences in the effect of climate warming on elevational shifts of harvesting sites
There were several significant interactions between climatic effects and geographical regions (Figure 6, for an overview of all model coefficients, see Supplemental Material S9). The hunting elevation of the rock ptarmigan was positively influenced by the mean ambient temperature in October and November and the annual fresh snow depth, with both effects more marked in the central and southern regions (Figure 6a,b). Hunting elevation of black grouse was positively affected by the number of frost days in October and November, the mean ambient temperature in October and November and the annual fresh snow depth, and all effects were most pronounced in the northern region (Figure 6c,d,e). In both hare species, we found no significant regional differences in the effect of climate warming on elevational shifts of harvesting sites (p > .05).
FIGURE 6.
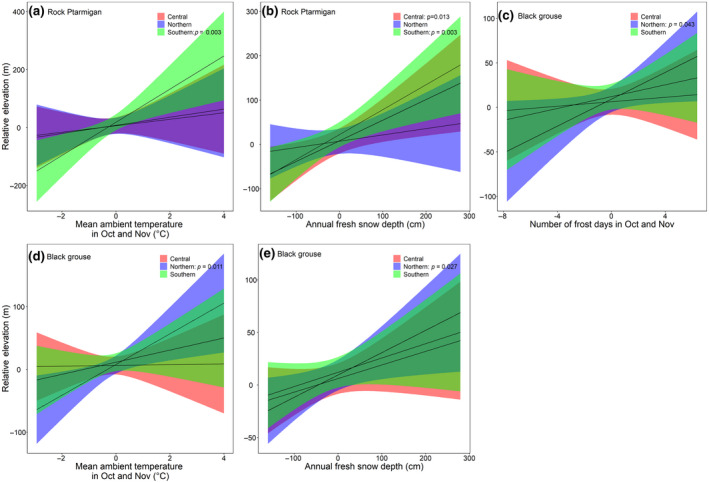
Relationships between different climate variables and the hunting elevation (m a.s.l.) of the four species collected over a 30‐year period (1991–2020) during the yearly hunting period in autumn in Grisons in the different biogeographical regions. Both hunting elevation and climate variables were detrended by yearly differencing. Effect of yearly differences of (a) mean ambient temperature in October and November (°C) and (b) annual fresh snow depth (cm) on relative elevation in rock ptarmigan, (c) number of frost days in October and November, (d) mean ambient temperature in October and November (°C) and (e) annual fresh snow depth (cm) on elevation in black grouse. The lines represent the regression lines with the red, blue and green areas showing the confidence intervals for the regions, respectively. Modelled on annual means of the data
4. DISCUSSION
4.1. Climate change in the mountains of Switzerland
Of the seven climate variables measured above 1200 m a.s.l., three variables (mean annual ambient temperature, mean ambient temperature in October and November and annual number of frost days) significantly changed within the 30 study years in the canton of Grisons. Accordingly, only these three climate variables showed climate warming in the Alpine environment of Grisons within the three decades of our study period. We were especially interested in climate variables describing snow occurrence as our study species included two alpine habitat specialists with coat colour change. None of the snow‐related climate variables changed within the three‐decade time period. Nevertheless, snow‐related climate variables in the Alps have previously been shown to be affected by climate warming but over longer time periods (>50 years; Marty & Blanchet, 2012; Nicolet et al., 2016). We suppose that our study period was not long enough to uncover the effect of climate change on annual snow depth, annual fresh snow depth and annual number of days with snow reliability in the Alps of Grisons. However, snow quality might be as important as snow quantity for species adapted to cold and snowy conditions.
4.2. Elevational shift in harvesting elevation
Contrary to our hypothesis, only the two grouse species clearly shifted their ranges towards higher elevations during the study period. Both shift rates exceed expected figures (Chen et al., 2011: median rate of 6.1–11.0 m decade−1) by factors of 3 (rock ptarmigan) and 5 (black grouse). If focussing on the minimum hunting elevation, the shift uphill was detectable in all four species, and in the two grouse species it was even more pronounced than the mean range shift. Minimum hunting elevation shifts increased more rapidly in tetraonids than in hares. Notably, the black grouse's large shift of minimum elevation cannot be attributed to a shift in the tree line, as effects of climate change on the tree line have not yet been detected in cultivated montane landscapes with artificially lowered tree lines, such as Grisons (Körner, 2003). If anything, Alpine forests have become thicker due to recent changes in forestry practices (Körner, 2003) which disadvantage this tetraonid. On the other hand, tetraonid species such as black grouse and rock ptarmigan use snow burrows to survive harsh winter conditions (Höhn, 2004; Klaus, 1990). To create burrows, they need dry powder snow (Klaus, 1990) which becomes scarce at lower altitudes due to global climate change. Hence, black grouse may find more suitable habitats at higher elevations as a consequence of increasing temperatures. In addition to that, rock ptarmigans have an even more limited range due to their adaptations (e.g. white winter plumage, which reduces predation risk in winter above the snow line, but increases it below the snow line). This bird species had the second highest shift rate and is likely to experience declines in habitat availability and increasing habitat fragmentation as warming continues. The effect of the time course on the maximum hunting elevation varied greatly between tetraonids and hares. Maximum hunting elevation in the Alpine mountain hare did not significantly change, whereas maximum hunting elevation in the European hare even decreased. This might probably be explained by the fact that hunters found game in the ‘traditional’ hunting areas and had not much reason to climb the mountains further up for hunting. In the tetraonids, the maximum hunting elevation increased considerably but the shift was less than in the overall hunting elevation. Altogether, the linear trend in the maximum hunting elevation was less clear as there was a larger variation between the years than in the minimum hunting elevation. An explanation might be that hunters adjusted their climbing activity to snow occurrence which was highly variable between the years. To sum up the results, tetraonids seem to respond more distinctively to climate warming than lagomorphs. Note that the R 2‐values of the models were consistently low because there was a high variance of hunting elevation per year.
4.3. Differences of elevational shifts in habitat specialists and generalists
In the tetraonids, hunting elevation of the habitat generalist (black grouse) increased more than in the habitat specialist (rock ptarmigan) during the three decades. The difference in shift to higher elevations was pronounced as the habitat generalist's ascent was almost twice faster than the habitat specialist's ascension. In all four species, the velocity of elevational shift was still higher at the trailing edge of habitat generalists’ (black grouse, European hare) in comparison to habitat specialists’ (rock ptarmigan, Alpine mountain hare) when examining the minimum hunting elevation. For both habitat generalist and specialist, the habitat requirements at the trailing edge may not be satisfied anymore due to climate warming and a rapid shift uphill is in progress. While the rapid shift uphill on the trailing edge can be almost maintained at the same velocity on the whole population range in the grouse species generalist, the rapid shift uphill on the trailing edge in the grouse species specialist is heavily slowed down over the whole population range. This implies that competition between the two grouse species for suitable habitat might increase with ongoing climate warming. Although the mean elevation of both hare species did not significantly change within the study period, the minimum hunting elevation shifted uphill and more rapidly in the European hare than in the Alpine mountain hare. This is an indication that the European hare might displace the Alpine mountain hare from its traditional range in the Alps due to global climate change. Our results are in line with several models and projections under global climate change illustrating the scenario that European hares have the potential to expand into mountain hare territory and that, after a time of sympatry due to range expansion by the European hare, the mountain hare disappears from optimal European hare habitat (Bisi et al., 2015; Caravaggi et al., 2017; La Morgia & Venturino, 2017; Leach, Kelly, et al., 2015, Leach, Montgomery, et al., 2015; Leach et al., 2017). As well as spatial displacement, introgressive hybridization poses a threat to the Alpine mountain hare by reducing its genetic integrity. Hybridization has been confirmed in hares from Grisons (Zachos et al., 2010). Based on our results, we predict that the rate of hybridization in the Alps will increase in the future. Our findings support the postulation that especially at high elevations, where the effects of climate warming are pronounced, climate warming leads to changes in species’ ranges (Chen et al., 2011). Within the order Lagomorpha, it has been shown that closely related species of similar size occurring at high elevations are likely to exhibit competitive interactions (Leach, Montgomery, et al., 2015), and that habitat generalists are less likely to be threatened by extinction than habitat specialists (Verde Arregoitia et al., 2015). This suggests that interacting species at high elevations, such as the European hare and the Alpine mountain hare, are likely to be highly susceptible to future environmental change. Our results covering only the last three decades indicate that the two lagomorph species already change their spatial distribution in the Alps.
4.4. Effect of climate variables on elevational shifts of harvesting sites
We detrended our data when analysing time series to assess whether there are not only trends over time but also a causal relationship between climate and elevation. In both tetraonids, annual fresh snow depth and number of frost days had an effect on elevational shifts in harvesting site. However, only the effect of the annual number of frost days on the elevational shifts of the black grouse was caused by climate warming as the other climate variables did not change significantly within the study years. Nevertheless, an increase in annual snow depth in both grouse species increased the hunting elevation. The use of snow burrows by both tetraonid species to survive harsh winter conditions (Höhn, 2004; Klaus, 1990) may explain this correlation. As snow‐related climate variables in the Alps have been shown to be affected by climate warming but over longer time periods, our results give indications that global climate change will have an impact on the two grouse species’ elevational range in the future. In the Alpine mountain hare, the number of frost days at the period of the hunt and the mean annual ambient temperature were influencing the hunting elevation. Only the effect of the mean annual ambient temperature on the hunting elevations of the Alpine mountain hare was caused by climate warming as the number of frost days in October and November did not change significantly within the study years. Unexpectedly, the rock ptarmigan and the Alpine mountain hare showed a decrease in relative hunting elevations when number of frost days in October and November or mean annual ambient temperature increased. The rock ptarmigan's decrease was not caused by climate warming and might be explained by feeding habits of this grouse species. As the rock ptarmigan feeds on different plant parts (Höhn, 2004), in autumns characterized by a high number of frost days better and more food might be available at lower elevations. However, the decrease of the Alpine mountain hare elevation was related to climate change as mean annual ambient temperature changed significantly within the study years. Not only changes this result the view on how snow‐adapted species react to climate change completely but also enhances the unpredictability of the future for snow‐adapted species under global climate change. European hares were not affected significantly by any climate variable. It seems as if this generalist species copes very well with different climate conditions within its elevation range.
4.5. Effect of biogeographical region on elevational shifts of harvesting sites
Hunting elevation of all four species in the Alps increased from the northern to the central to the southern biogeographical region of Grisons. This corresponds well to the main climatic difference that winters become milder from northern to southern Grisons (MeteoSchweiz, 2012). Alpine mountain hares showed a significant shift to higher elevations in the northern and southern biogeographical region. The shift rates did not differ between these two biogeographical regions. In comparison, no significant elevational shift in the European hare was recorded in the three biogeographical regions. In tetraonids, the shift uphill was especially steep in the southern biogeographical region of Grisons. Moreover, while the black grouse ascended significantly in the northern biogeographical region of Grisons, for the rock ptarmigan no significant increment was recorded. Our results display that the different ecological requirements of the involved species result in a highly variable and, consequently, unpredictable outcome how species respond to recent warming.
4.6. Regional differences in the effect of climate warming on elevational shifts of harvesting sites
In the black grouse, the elevational shifts of harvesting sites were influenced by climatic variables in the northern region only. In rock ptarmigans, the elevational shifts were influenced by climatic variables in the central and southern region. Both species were affected by either mean annual ambient temperature or mean ambient temperature in October and November which both showed significant changes during the last decades in Grisons. Our findings show that the response of these two species to recent climate warming is not only affected by complex interactions between biogeographical region and climate variable, but also differ with respect to their slope (positive or negative). This underlies that the outcome of how species respond to recent warming can be difficult to predict locally. The hunting elevation of both hare species was not influenced by any interaction between climate variables and biogeographical region during the last three decades.
4.7. Wildlife management in the canton of Grisons
Grisons has a mean elevation of 2027 m a.s.l. (Meier & Schmid, 2007) and is the political unit in the Alps with the highest mean elevation. Around 50% of the canton’s surface lies above the timberline in the alpine and nival zone. In this respect, Grisons is highly different from the neighbouring cantons and countries. Even a major rise in the timberline will change the extent of the extensive alpine habitats only slightly in most regions of Grisons. Therefore, this canton has buffer capacities against the negative effects of climate warming on Alpine species adapted to cold and snowy conditions like hardly any other region. It might be possible that this is also the reason that within the three decades covered by our study in both hare species less elevational range shift was recorded as we initially had expected. Nevertheless, it is important in Grisons also to document the consequences of climate warming and take these changes into account in the wildlife management of these four hunted species. Due to the monitoring of the four species, the game sanctuaries, and the game refuges in winter, it is not surprising that the conservation status of all four species in Grisons differs from that in other alpine regions. All four species in Grisons have high population densities that benefit from the large occurrence of alpine habitats, from cautious wildlife management and from the large interest by the public.
4.8. Importance of wildlife monitoring and management under global climate change
We used a large, long‐term dataset to demonstrate that, for birds and mammal species adapted to cold conditions, the rate of range shift in the wake of climate warming is smaller than for congeneric generalist species. Hence, all four taxa respond to climate change but habitat generalists more rapidly than habitat specialists. The available surface area decreases as animal species shift uphill. Thus, habitat specialists may lose habitat availability more rapidly than habitat generalists. As warming continues in future, the elevation of the tree line will increase slowly (Körner, 2003; Lu et al., 2021). The elevation of the snow line will increase rapidly (Beniston et al., 2003), resulting in reduced winter habitat and increased habitat fragmentation for alpine species, especially in the lower Alps. Climate change is thus demonstrated to form a serious threat especially to alpine biodiversity. Consequently, regions rich in subalpine and alpine habitats will have an increased responsibility to conserve these species, for example, by implementing wildlife monitoring and sustainable hunting strategies like in Grisons. As Alpine mammals and birds are appropriate flagship species, our results make a strong contribution to the urgent call to disseminate information on the detrimental consequences of climate change.
CONFLICT OF INTEREST
The author declares no conflict of interest.
Supporting information
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
ACKNOWLEDGEMENTS
We are grateful to all hunters of Grison as licence holders as well as all collaborators of the Amt für Jagd und Fischerei Graubünden (game wardens, hunters, etc.) for providing data on hunting elevations. Climate data were provided by MeteoSwiss.
Schai‐Braun, S. C. , Jenny, H. , Ruf, T. , & Hackländer, K. (2021). Temperature increase and frost decrease driving upslope elevational range shifts in Alpine grouse and hares. Global Change Biology, 27, 6602–6614. 10.1111/gcb.15909
DATA AVAILABILITY STATEMENT
Data available in article supplementary material (S1 Table, S2 Table, S3 Table, S5 Table).
REFERENCES
- Averianov, A. , Niethammer, J. , & Pegel, M. (2003). Lepus europaeus Pallas, 1778—Feldhase. In Niethammer J. & Krapp F. (Eds.), Handbuch der Säugetiere Europas, Band 3/II Hasentiere (pp. 35–104). AULA‐Verlag. [Google Scholar]
- Balzari, C. , & Gygax, A. (2019). Vogelarten der Schweiz. Der Bestimmungsführer. 2., überarbeitete und erweiterte Auflage 2019. Haupt Verlag. [Google Scholar]
- Bartoń, K. (2013). MuMIn: Multi‐model inference. R package version 1.9.13. http://CRAN.R‐project.org/package=MuMIn
- Bellard, C. , Bertelsmeier, C. , Leadley, P. , Thuiller, W. , & Courchamp, F. (2012). Impacts of climate change on the future of biodiversity. Ecology Letters, 15(4), 365–377. 10.1111/j.1461-0248.2011.01736.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beniston, M. , Keller, F. , Koffi, B. , & Goyette, S. (2003). Estimates of snow accumulation and volume in the Swiss Alps under changing climatic conditions. Theoretical and Applied Climatology, 76(3–4), 125–140. 10.1007/s00704-003-0016-5 [DOI] [Google Scholar]
- Bisi, F. , Wauters, L. , Preatoni, D. , & Martinoli, A. (2015). Interspecific competition mediated by climate change: Which interaction between brown and mountain hare in the Alps? Mammalian Biology, 80(5), 424–430. 10.1016/j.mambio.2015.06.002 [DOI] [Google Scholar]
- Büntgen, U. , Greuter, L. , Bollmann, K. , Jenny, H. , Liebhold, A. , Galván, J. , Stenseth, N. , Andrew, C. , & Mysterud, A. (2017). Elevational range shifts in four mountain ungulate species from the Swiss Alps. Ecosphere, 8, 4. [Google Scholar]
- Caravaggi, A. , Leach, K. , Santilli, F. , Rintala, J. , Helle, P. , Tiainen, J. , Bisi, F. , Martinoli, A. , Montgomery, W. , & Reid, N. (2017). Niche overlap of mountain hare subspecies and the vulnerability of their ranges to invasion by the European hare; the (bad) luck of the Irish. Biological Invasions, 19(2), 655–674. 10.1007/s10530-016-1330-z [DOI] [Google Scholar]
- Chen, I.‐C. , Hill, J. , Ohlemüller, R. , Roy, D. , & Thomas, C. (2011). Rapid range shifts of species associated with high levels of climate warming. Science, 333, 1024–1026. 10.1126/science.1206432 [DOI] [PubMed] [Google Scholar]
- Faraway, J. (2006). Extending the linear model with R. Texts in statistical science. Chapman and Hall/CRC. [Google Scholar]
- Feldmeier, S. , Schmidt, B. , Zimmermann, N. , Veith, M. , Ficetola, G. , & Lötters, S. (2020). Shifting aspect or elevation? The climate change response of ectotherms in a complex mountain topography. Diversity and Distributions, 26(11), 1483–1495. 10.1111/ddi.13146 [DOI] [Google Scholar]
- Fox, J. , & Weisberg, S. (2019). An R companion to applied regression (3rd ed.). Sage. [Google Scholar]
- Gidoin, C. , Roques, L. , & Boivin, T. (2015). Linking niche theory to ecological impacts of successful invaders: Insights from resource fluctuation‐specialist herbivore interactions. Journal of Animal Ecology, 84, 396–406. 10.1111/1365-2656.12303 [DOI] [PubMed] [Google Scholar]
- Gonseth, Y. , Wohlgemuth, T. , Sansonnens, B. , & Butler, A. (2001). Die biogeographischen Regionen der Schweiz. Erläuterungen und Einteilungsstandard. Umwelt‐Materialien Nr. 137. Wald und Landschaft BUWAL Bundesamt für Umwelt, Berne.
- Gottfried, M. , Pauli, H. , Futschik, A. , Akhalkatsi, M. , Barančok, P. , Benito Alonso, J. , Coldea, G. , Dick, J. , Erschbamer, B. , Fernández Calzado, M. , Kazakis, G. , Krajči, J. , Larsson, P. , Mallaun, M. , Michelsen, O. , Moiseev, D. , Moiseev, P. , Molau, U. , Merzouki, A. , … Grabherr, G. (2012). Continent‐wide response of mountain vegetation to climate change. Nature Climate Change, 2(2), 111–115. 10.1038/nclimate1329 [DOI] [Google Scholar]
- Hackländer, K. , & Schai‐Braun, S. C. (2018). Lepus europaeus. In Smith A. T., Johnston C. H., Alves P. C., & Hackländer K. (Eds.), Lagomorphs—Pikas, rabbits, and hares of the world. Johns Hopkins University Press. [Google Scholar]
- Harsch, M. , Hulme, P. , McGlone, M. , & Duncan, R. (2009). Are treelines advancing? A global meta‐analysis of treeline response to climate warming. Ecology Letters, 12(10), 1040–1049. 10.1111/j.1461-0248.2009.01355.x [DOI] [PubMed] [Google Scholar]
- Höhn, O. (2004). Die Schneehühner. Gattung Lagopus. Westarp Wiss.‐Verl.‐Ges, Hohenwarsleben, Die neue Brehm‐Bücherei, Vol. 408.
- Hothorn, T. , Bretz, F. , & Westfall, P. (2008). Simultaneous inference in general parametric models. Biometrical Journal, 503, 346–363. 10.1002/bimj.200810425 [DOI] [PubMed] [Google Scholar]
- IPCC . (2014). Climate change 2014: Impacts, adaptation and vulnerability. In Field C. B., Barros V. R., Dokken D. J., Mach K. J., Mastrandrea M. D., Bilir T. E., Chatterjee M., Ebi K. L., Estrada Y. O., Genova R. C., Girma B., Kissel E. S., Levy A. N., MacCracken S., Mastrandrea P. R., & White L. L. (Eds.), Contribution of working group II to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press. [Google Scholar]
- IPCC . (2021). Climate change 2021: The physical science basis. Contribution of working group I to the sixth assessment report of the Intergovernmental Panel on Climate Change. In Masson‐Delmotte V., Zhai P., Pirani A., Connors S. L., Péan C., Berger S., Caud N., Chen Y., Goldfarb L., Gomis M. I., Huang M., Leitzell K., Lonnoy E., Matthews J. B. R., Maycock T. K., Waterfield T., Yelekçi O., Yu R., & Zhou B. (Eds.). Cambridge University Press. Cambridge. [Google Scholar]
- Johnson, D. , Büntgen, U. , Frank, D. , Kausrud, K. , Haynes, K. , Liebhold, A. , Esper, J. , & Stenseth, N. (2010). Climatic warming disrupts recurrent Alpine insect outbreaks. Proceedings of the National Academy of Sciences of the United States of America, 107(47), 20576–20581. 10.1073/pnas.1010270107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Just, M. (2013). Material zum Monitoring des Alpenschneehuhns (Lagopus muta) im Kanton Graubünden. Amt für Jagd und Fischerei Graubünden. [Google Scholar]
- Klaus, S. (1990). Die Birkhühner. Tetrao tetrix und T. mlokosiewiczi. Die neue Brehm‐Bücherei (Vol. 397). VerlagsKG Wolf. [Google Scholar]
- Körner, C. (2003). Alpine plant life. Functional plant ecology of high mountain ecosystems (2nd ed.). Springer. [Google Scholar]
- La Morgia, V. , & Venturino, E. (2017). Understanding hybridization and competition processes between hare species: Implications for conservation and management on the basis of a mathematical model. Ecological Modelling, 364, 13–24. 10.1016/j.ecolmodel.2017.09.002 [DOI] [Google Scholar]
- Langbein, J. , Hutchings, M. R. , Harris, S. , Stoate, C. , Tapper, S. C. , & Wray, S. (1999). Techniques for assessing the abundance of brown hares Lepus europaeus . Mammal Review, 29(2), 93–116. 10.1046/j.1365-2907.1999.00040.x [DOI] [Google Scholar]
- Leach, K. , Kelly, R. , Cameron, A. , Montgomery, W. , & Reid, N. (2015). Expertly validated models and phylogenetically‐controlled analysis suggests responses to climate change are related to species traits in the order Lagomorpha. PLoS ONE, 10(4), e0122267. 10.1371/journal.pone.0122267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leach, K. , Montgomery, W. , & Reid, N. (2015). Biogeography, macroecology and species’ traits mediate competitive interactions in the order Lagomorpha. Mammal Review, 45, 88–102. 10.1111/mam.12035 [DOI] [Google Scholar]
- Leach, K. , Montgomery, W. , & Reid, N. (2017). Characterizing biotic interactions within the Order Lagomorpha using Joint Species Distribution Models at 3 different spatial scales. Journal of Mammalogy, 18, 1509. 10.1093/jmammal/gyx105 [DOI] [Google Scholar]
- Lenoir, J. , Gégout, J. , Marquet, P. , de Ruffray, P. , & Brisse, H. (2008). A significant upward shift in plant species optimum elevation during the 20th century. Science, 320(5884), 1768–1771. 10.1126/science.1156831 [DOI] [PubMed] [Google Scholar]
- Loarie, S. , Duffy, P. , Hamilton, H. , Asner, G. , Field, C. , & Ackerly, D. (2009). The velocity of climate change. Nature, 462(7276), 1052–1055. 10.1038/nature08649 [DOI] [PubMed] [Google Scholar]
- Lu, X. , Liang, E. , Wang, Y. , Babst, F. , & Camarero, J. (2021). Mountain treelines climb slowly despite rapid climate warming. Global Ecology and Biogeography, 30(1), 305–315. 10.1111/geb.13214 [DOI] [Google Scholar]
- Marty, C. , & Blanchet, J. (2012). Long‐term changes in annual maximum snow depth and snowfall in Switzerland based on extreme value statistics. Climatic Change, 111(3–4), 705–721. 10.1007/s10584-011-0159-9 [DOI] [Google Scholar]
- Meier, C. , & Schmid, H. (2007). Die Vögel Graubündens. 3., vollst. überarb. Aufl. Verlag Desertina, Chur, Graubünden Books. [Google Scholar]
- MeteoSchweiz . (2012). Klimabericht Kanton Graubünden 2012. Bundesamt für Meteorologie und Klimatologie MeteoSchweiz. Amtes für Natur und Umwelt Graubünden. (Fachbericht MeteoSchweiz, 242, 242).
- Nicolet, G. , Eckert, N. , Morin, S. , & Blanchet, J. (2016). Decreasing spatial dependence in extreme snowfall in the French Alps since 1958 under climate change. Journal of Geophysical Research: Atmospheres, 121(14), 8297–8310. 10.1002/2016JD025427 [DOI] [Google Scholar]
- Parmesan, C. , & Yohe, G. (2003). A globally coherent fingerprint of climate change impacts across natural systems. Nature, 421(6918), 37–42. 10.1038/nature01286 [DOI] [PubMed] [Google Scholar]
- Pauli, H. , Gottfried, M. , Dullinger, S. , Abdaladze, O. , Akhalkatsi, M. , Benito Alonso, J. , Coldea, G. , Dick, J. , Erschbamer, B. , Fernández Calzado, R. , Ghosn, D. , Holten, J. , Kanka, R. , Kazakis, G. , Kollár, J. , Larsson, P. , Moiseev, P. , Moiseev, D. , Molau, U. , … Grabherr, G. (2012). Recent plant diversity changes on Europe's mountain summits. Science, 336(6079), 353–355. 10.1126/science.1219033 [DOI] [PubMed] [Google Scholar]
- Post, E. , Forchhammer, M. , Bret‐Harte, M. , Callaghan, T. , Christensen, T. , Elberling, B. , Fox, A. , Gilg, O. , Hik, D. , Høye, T. , Ims, R. , Jeppesen, E. , Klein, D. , Madsen, J. , McGuire, A. , Rysgaard, S. , Schindler, D. , Stirling, I. , Tamstorf, M. , … Aastrup, P. (2009). Ecological dynamics across the Arctic associated with recent climate change. Science, 325(5946), 1355–1358. 10.1126/science.1173113 [DOI] [PubMed] [Google Scholar]
- R Development Core Team . (2019). R: A language and environment for statistical computing. R Foundation for Statistical Computing. www.R‐project.org [Google Scholar]
- Reid, N. , Brommer, J. , Stenseth, N. , Marnell, F. , McDonald, R. , & Montgomery, W. (2021). Regime shift tipping point in hare population collapse associated with climatic and agricultural change during the very early 20th century. Global Change Biology, 15(05), 3732–3740. 10.1111/gcb.15652 [DOI] [PubMed] [Google Scholar]
- Reif, J. , & Flousek, J. (2012). The role of species’ ecological traits in climatically driven altitudinal range shifts of central European birds. Oikos, 121(7), 1053–1060. 10.1111/j.1600-0706.2011.20008.x [DOI] [Google Scholar]
- Schenker, L. , Bollmann, K. , Rehnus, M. , Brodbeck, S. , & Gugerli, F. (2020). Hare's affairs: Lessons learnt from a noninvasive genetic monitoring for tracking mountain hare individuals. Ecology and Evolution, 10(18), 10150–10166. 10.1002/ece3.6676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas, C. , Cameron, A. , Green, R. , Bakkenes, M. , Beaumont, L. , Collingham, Y. , Erasmus, B. N. , Ferreira de Siqueira, M. , Grainger, A. , Hannah, L. , Hughes, L. , Huntley, B. , van Jaarsveld, A. , Midgley, G. , Miles, L. , Ortega‐Huerta, M. , Townsend Peterson, A. , Phillips, O. L. , & Williams, S. (2004). Extinction risk from climate change. Nature, 427, 145–148. 10.1038/nature02121 [DOI] [PubMed] [Google Scholar]
- van Rij, J. , Wieling, M. , Baayen, R. , & van Rijn, H. (2020). itsadug: Interpreting time series and autocorrelated data using GAMMs.
- Verde Arregoitia, L. , Leach, K. , Reid, N. , & Fisher, D. (2015). Diversity, extinction, and threat status in Lagomorphs. Ecography, 38, 1155–1165. 10.1111/ecog.01063 [DOI] [Google Scholar]
- Wood, S. (2004). Stable and efficient multiple smoothing parameter estimation for generalized additive models. Journal of the American Statistical Association, 99(467), 673–686. 10.1198/016214504000000980 [DOI] [Google Scholar]
- Zachos, F. , Slimen, H. , Hackländer, K. , Giacometti, M. , & Suchentrunk, F. (2010). Regional genetic in situ differentiation despite phylogenetic heterogeneity in Alpine mountain hares. Journal of Zoology, 282, 47–53. 10.1111/j.1469-7998.2010.00710.x [DOI] [Google Scholar]
- Zuur, A. , Ieno, E. , Walker, N. , Saveliev, A. , & Smith, G. (2009). Mixed effects models and extensions in ecology with R. Springer. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Data Availability Statement
Data available in article supplementary material (S1 Table, S2 Table, S3 Table, S5 Table).