
Fig. 1.
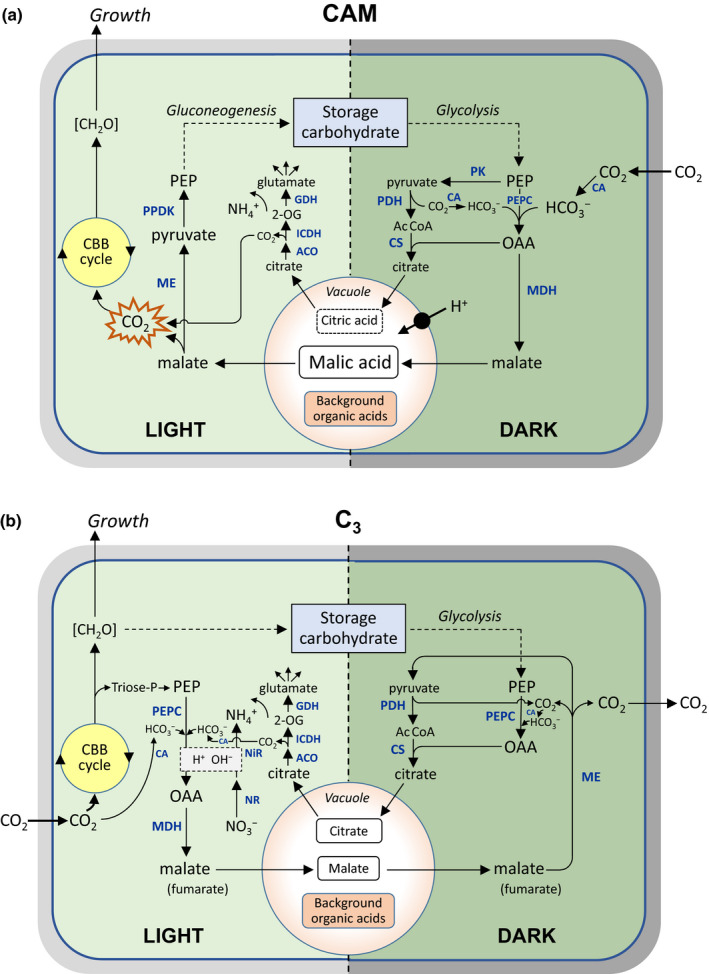
Principal day–night carbon fluxes and associated carboxylate metabolism in photosynthetic mesophyll cells, highlighting similarities and differences between crassulacean acid metabolism (CAM) plants and C3 plants. The schemes are modelled on the format used by Schiller & Bräutigam (2021) and show steady‐state operation of the cycles over the 24 h period. For simplicity, details of the energetics of the pathways, subcellular compartmentation and charges (dissociation states) of the metabolites have been omitted, but further information can be found in Winter & Smith (1996a); Holtum et al. (2005) and Shameer et al. (2018). Additional internal sources of CO2, such as the tricarboxylic acid (TCA) cycle, oxidative pentose phosphate pathway and photorespiration, are not shown explicitly. The dashed lines to and from storage carbohydrate indicate multi‐step pathways. The central vacuole is shown schematically, but in mature mesophyll cells can typically occupy ≥ 90% of the cell volume. (a) In CAM plants, net nocturnal CO2 fixation via phosphoenolpyruvate (PEP) carboxylase (PEPC) results in vacuolar accumulation of malic acid (2 H+ per malate); this is mobilized in the following daytime and decarboxylated (either via NAD(P)‐ME, as shown, or in some CAM plants via PEP carboxykinase), with the CO2 released behind closed stomata at high concentration being refixed by Rubisco in the Calvin–Benson–Bassham (CBB) cycle. In some CAM plants there is significant nocturnal synthesis and vacuolar accumulation of citric acid: this is not associated with net fixation of CO2, but on remobilization in the following daytime, the citrate supplies carbon skeletons for assimilation via glutamate dehydrogenase (GDH) and glutamine synthetase (the latter not shown explicitly). Nocturnal acid accumulation requires net influx of H+ into the vacuole, which is driven by the tonoplast H+ pump(s) (see main text). The vacuole also contains a background pool of organic acids that does not oscillate during the day–night cycle. Carbon skeletons required to support nocturnal acid accumulation are provided by glycolytic breakdown of storage carbohydrate (usually chloroplastic starch/glucan, but significant vacuolar hexose in some species), which is regenerated by gluconeogenesis in the light period. (b) In C3 plants, citrate synthesis and vacuolar accumulation also occur at night, but malate accumulation (and fumarate in some species) typically occurs during the daytime, which provides charge‐ and pH‐balancing for light‐dependent nitrate assimilation (light grey box). In some plants such as Arabidopsis thaliana, citrate and malate fluxes can be of similar magnitude, as shown here to achieve flux balance across the vacuolar membrane, but diel carboxylate accumulation and remobilization are not charge‐balanced by H+ and so do not result in vacuolar pH changes during the day–night cycle (see Table 1). AcCoA, acetyl‐CoA; ACO, aconitase; CA, carbonic anhydrase; [CH2O], sugars/carbohydrate; CS, citrate synthase; ICDH, isocitrate dehydrogenase; MDH, malate dehydrogenase; ME, malic enzyme; NiR, nitrite reductase; NR, nitrate reductase; OAA, oxaloacetate; 2‐OG, 2‐oxoglutarate; PDH, pyruvate dehydrogenase; PK, pyruvate kinase; PPDK, pyruvate, Pi dikinase.