

Abstract
CRISPR-based “active genetic” elements developed in 2015 bypassed the fundamental rules of traditional genetics. Inherited in a super-Mendelian fashion, such selfish genetic entities offered a variety of potential applications including: gene-drives to disseminate gene cassettes carrying desired traits throughout insect populations to control disease vectors or pest species, allelic drives biasing inheritance of preferred allelic variants, neutralizing genetic elements to delete and replace or to halt the spread of gene-drives, split-drives with the core constituent Cas9 endonuclease and guide RNA (gRNA) components inserted at separate genomic locations to accelerate assembly of complex arrays of genetic traits or to gain genetic entry into novel organisms (vertebrates, plants, bacteria), and inter-homolog based copying systems in somatic cells to develop tools for treating inherited or infectious diseases. Here, we summarize the substantial advances that have been made on all of these fronts and look forward to the next phase of this rapidly expanding and impactful field.
Keywords: active genetics, gene-drive, allelic-drive, Pro-Active Genetics, inter-homolog pair, non-homologous end-joining, homology-directed repair, Drosophila
Introduction
In 2016 Valentino Gantz, then a graduate student in my lab, and I published a forward-looking review in this same journal [1] in which we proposed diverse hypothetical applications of a new field we termed “active genetics” (Fig. 1A). The prototype of these active genetic elements consisted of a gene cassette encoding the Cas9 endonuclease and a guide RNA (gRNA) that was inserted into the Drosophila genome at the precise location where the gRNA directed DNA cleavage by Cas9 [2]. When active in the germline, Cas9 combined with the gRNA to cleave the homologous chromosome in the corresponding position as the gene cassette, resulting in the homology directed repair (HDR) pathway copying the Cas9 + gRNA carrying element into the DNA break through a local directional gene conversion event (Fig. 1B). By this copying mechanism, powerful gene-drive systems (Fig. 1C) can be harnessed to disseminate linked traits throughout a population (Fig. 1D). These gene-drives could be used to spread beneficial genetic traits such as effector proteins preventing transmission of malarial parasites by mosquitoes (population modification), or alternatively be deployed as destructive trojan horses to reduce or eliminate pest populations (population suppression). As described further below, we also envisioned other potential designs of self-propagating active genetic elements in which the Cas9 and gRNA were provided in various alternative configurations (Fig. 1A, Fig. 2).
Figure 1: Active Genetics in the germline.
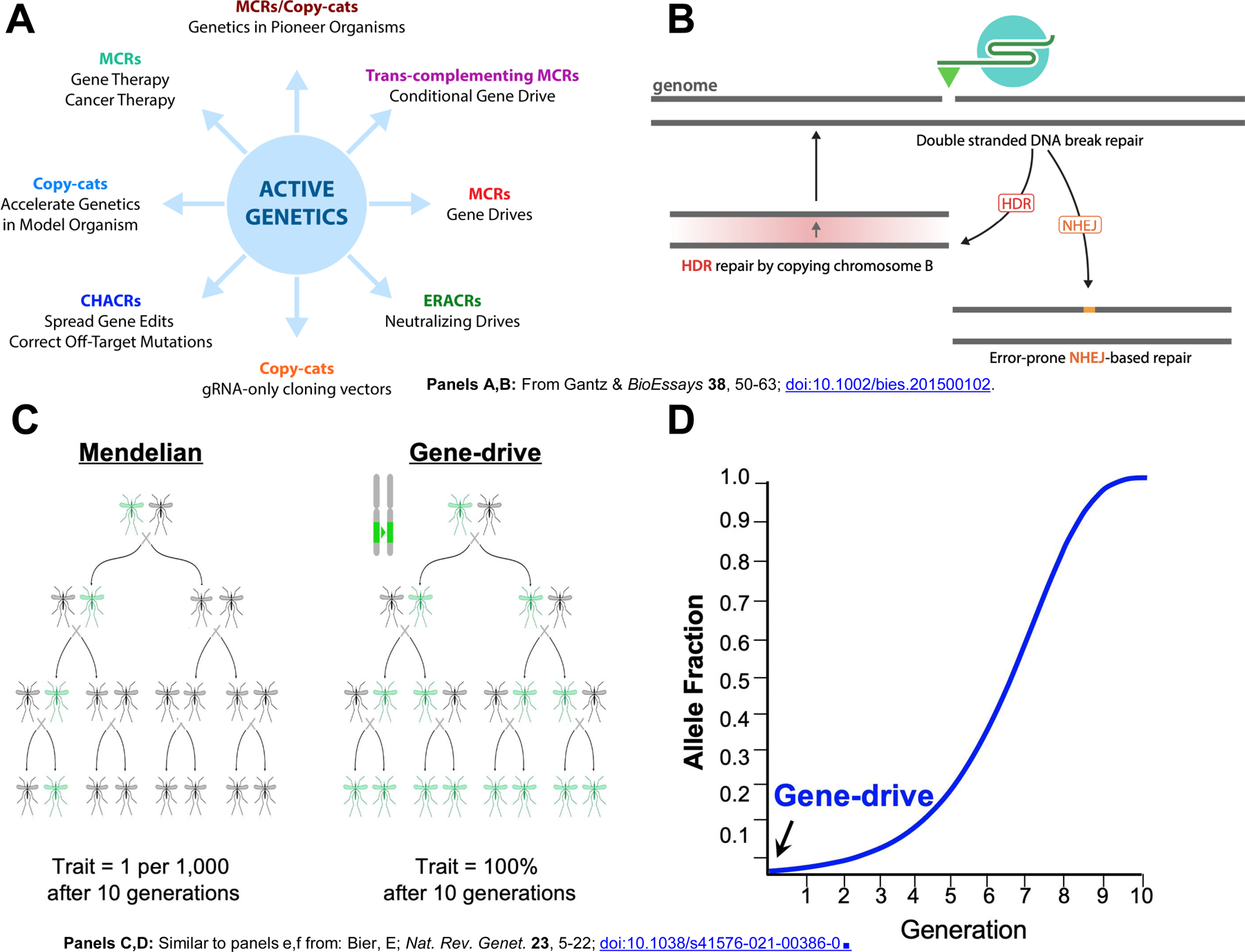
A) Reproduction of the original hypothetical wheel of possible active genetic applications proposed in the BioEssays review, the Dawn of Active Genetics, by Gantz and Bier, 2016 [1], which included all of the examples schematized in Fig. 2. (MCR = Mutagenic Chain Reaction, or original term for an active genetic element that copied in both germline and somatic cells [2]). B) Overview of double strand break (DSB) repair following Cas9/gRNA mediated cleavage of the genome highlighting precise homology dependent repair (HDR)versus potentially error prone non-homologous end-joining (NHEJ) (reproduced from [1]). C) Comparison of Mendelian versus “active genetics” or gene-drive inheritance over several generations. D) Example of predicted logistic growth profile for an ideal gene-drive over successive generations when introduced initially at a ratio of gene-drive:wild-type = 1:100.
Figure 2: Gene-drive systems.
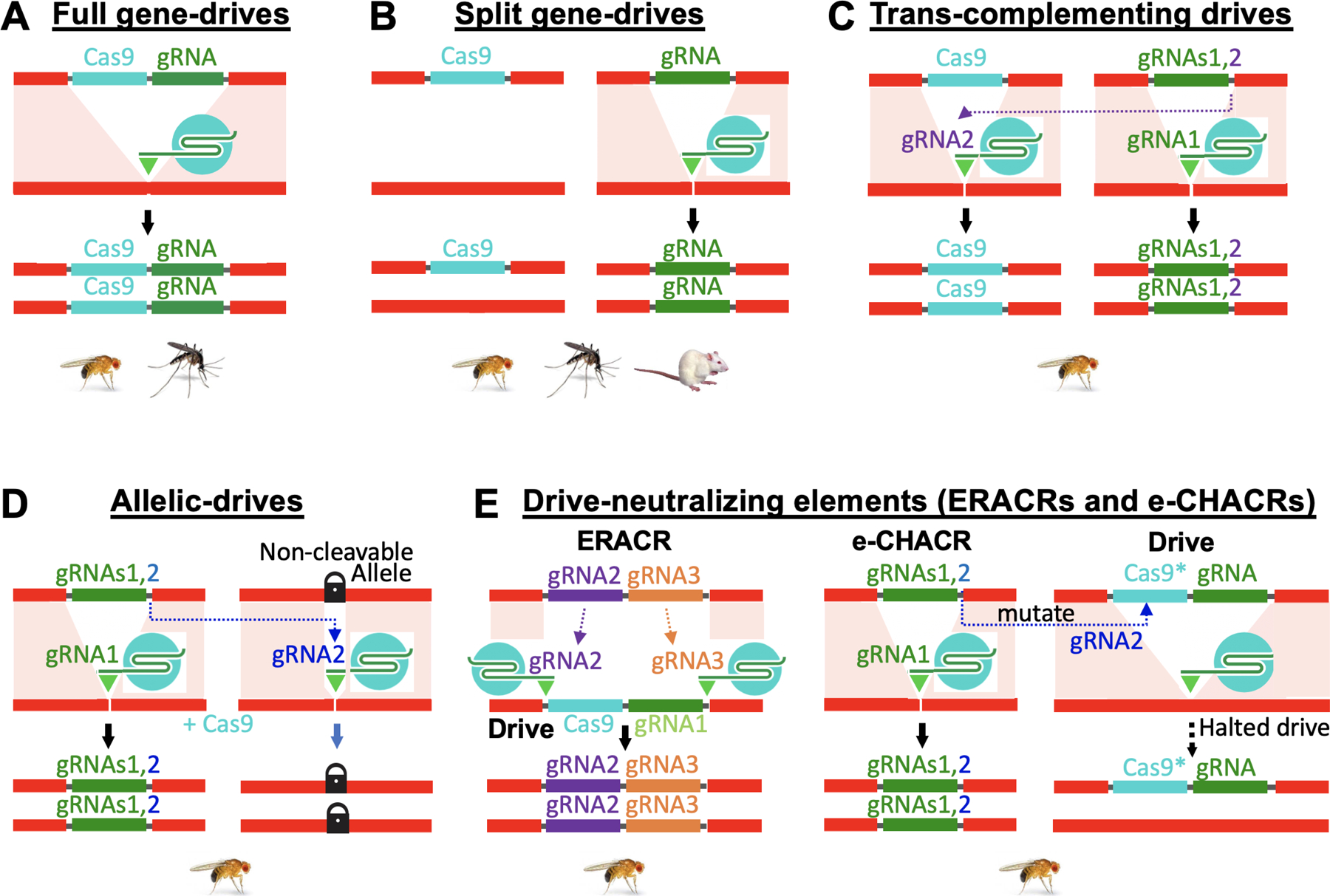
A) Diagram of a full gene drive in which the Cas9 and gRNA encoding transgenes are linked in a single cassette. The gRNA programs Cas9 to cleave the genome at the location where the cassette is inserted. Other genetic cargo such as anti-malarial effectors or additional gRNAs can also be carried and copied by the drive cassette [16]. B) A simple split-drive system in which the gRNA bearing cassette is inserted into the site where the gRNA directs Cas9 cleavage and the Cas9 transgene is inserted elsewhere in the genome where it is transmitted in a standard Mendelian fashion. When these two elements are combined, only the gRNA element is copied, leading to an additive accumulation of this drive component over multiple generations. The frequency of the Cas9 element will not change in such cases unless it carries a fitness cost either alone or in combination [38] with the gRNA element. C) A special category of split gene-drive in which the gRNA cassette carries an additional gRNA that directs cleavage at the insertion site of the Cas9 transgene. When this dual gRNA bearing element is co-inherited with the Cas9 element, the two gRNAs complex with Cas9 to copying both elements, creating a full co-drive configuration referred to as a trans-complementing drive [39]. D) Allelic drives carry a second gRNA, in addition to the one that directs its own copying, that cleaves the genome at a site present in an unfavored allelic variant but in not in the preferred allele. As the drive copies itself through a population (via gRNA1) it also copies the non-cleavable favored allele (via gRNA2) leading to increased frequencies of both the drive and favored allelic variant [18]. E) Drive-neutralizing elements, which carrying gRNAs but no Cas9 source, are designed either to delete and replace the drive (ERACRs) or to mutagenize Cas9 to halt the drive process (e-CHACRs). ERACRs (left) are inserted at the same genomic site as a gene-drive and encode two gRNAs that direct Cas9 (produced by the drive element) to cut on either side of the drive element to delete it. The resulting gap is then repaired by copying the ERACR which replaces the drive element. e-CHACRs (right) carry one gRNA to direct copying the element (at any site in the genome) and a second a gRNA targeting Cas9 for mutagenesis. When an e-CHACR confronts a drive element it copies itself and then mutates Cas9 to halt the drive process. In panels A-E, icons below the diagrams indicate the different organisms (flies, mosquitoes, or mice) in which the various drive systems have been developed thus far.
Background
Barbara McClintock, a legendary cytogeneticist and Nobel laureate, discovered mobile genetic elements, or transposons, in maize [3]. The selfish Dissociation (Ds) genetic elements McClintock studied can jump out of one genomic site and into another unlike traditional genes whose location remains stable and constant. The Ds element, like many mobile elements, transposes via a cut-and-paste mechanism leading to mottled mosaic pigmentation patterns of kernels [4,5]. Homing endonuclease genes (HEGs) [6,7] are another well-studied type of selfish genetic element in yeast that are transmitted to progeny at super-Mendelian frequencies (i.e., >50%). HEGS, which are similar to class-II sequence-specific restriction endonucleases, cleave extended, and thus very rare, recognition sequences and, critically, are inserted into the genome at their unique site of genome cleavage. When a diploid yeast cell is heterozygous for HEG and a wild-type chromosome, the HEG endonuclease cuts the opposing chromosome and copies itself into the break via the homology directed repair (HDR) pathway (Fig. 1B).
In addition to selfish genetic elements such as transposons or HEGs, entire chromosomes also can manifest non-Mendelian transmission or “drive”. For example, sex chromosomes often display a phenomenon referred to as meiotic drive in which either the male or female determining chromosome is preferentially transmitted to offspring [8–10]. Similarly, balanced translocations which are infertile in the trans-heterozygous state, were proposed by Chris Curtis in the 1960s as a means for spreading favorable genetic traits into a population [11].
The identification and molecular characterization of specific selfish genetic elements such as HEGs led mathematical modelers such as pioneer Austin Burt and colleagues to develop a modern framework for engineering HEG-based gene-drive systems to introduce deleterious genetic traits into populations that either lead to lethality or sterility when homozygous [12]. A significant limitation of HEG-based gene-drive systems, however, is that their target recognition sites cannot be readily altered [13–15]. The advent of efficient programmable CRISPR-based gene drives described below overcame this constraint, fueling great progress in this field and validating many of Burt’s visionary predictions.
The many facets of active genetics
As covered in our original review [1], co-author Valentino Gantz constructed and tested the first CRISPR-based meiotic gene-drive in fruit flies [2]. He then built a much larger complex construct for mosquitoes carrying dual anti-malarial effector genes. In collaboration with the laboratory of Anthony James (UC-Irvine), this construct was inserted into the kynurenine hydroxylase (kh) locus in mosquitoes (Anopheles stephensi). Transgenic males carrying this construct were found to transmit it to ~99% of their offspring, although as discussed below, this element did not drive as efficiently when transmitted by females [16].
In our previous review we also envisioned several configurations with unlinked Cas9 and gRNA components (split systems) that could be used for diverse purposes including neutralizing genetic elements designed either to delete and replace gene drives or to halt their spread, conditional trans-complementing drive systems, and locally confinable drive systems (Fig. 1A).
In this current follow-up review, we first survey the remarkable progress that has been made on all these fronts of active genetics. We begin by briefly discussing strategies to improve performance of gene-drives. Next, we summarize progress on developing allelic drive systems [17,18] to bias inheritance of preferred naturally occurring allelic variants, neutralizing drives that delete and replace a gene-drive [19], halt a drive by mutating the Cas9 endonuclease [19,20]), or inhibit Cas9 activity by expressing Cas9-inhibitory proteins [21] (Fig. 2). Circling back to the original observations in which the prototype gene-drive also generated highly penetrant somatic phenotypes, we summarize studies of genetic elements referred to as CopyCatchers designed to detect and quantify inter-homolog copying events in somatic cells [22]. We discuss these strategies in the context of other newly developed in vivo gene-editing systems, as well as generalizing active genetic systems to new organisms including vertebrates [23,24], plants, and bacteria [25] (e.g., to reduce the incidence of antibiotic resistance) (Figs. 3–5). Following each section, we also look forward to development of new and potentially transformative extensions of these various avenues of research (Fig. 6).
Figure 3: Active genetics in bacteria and viruses.
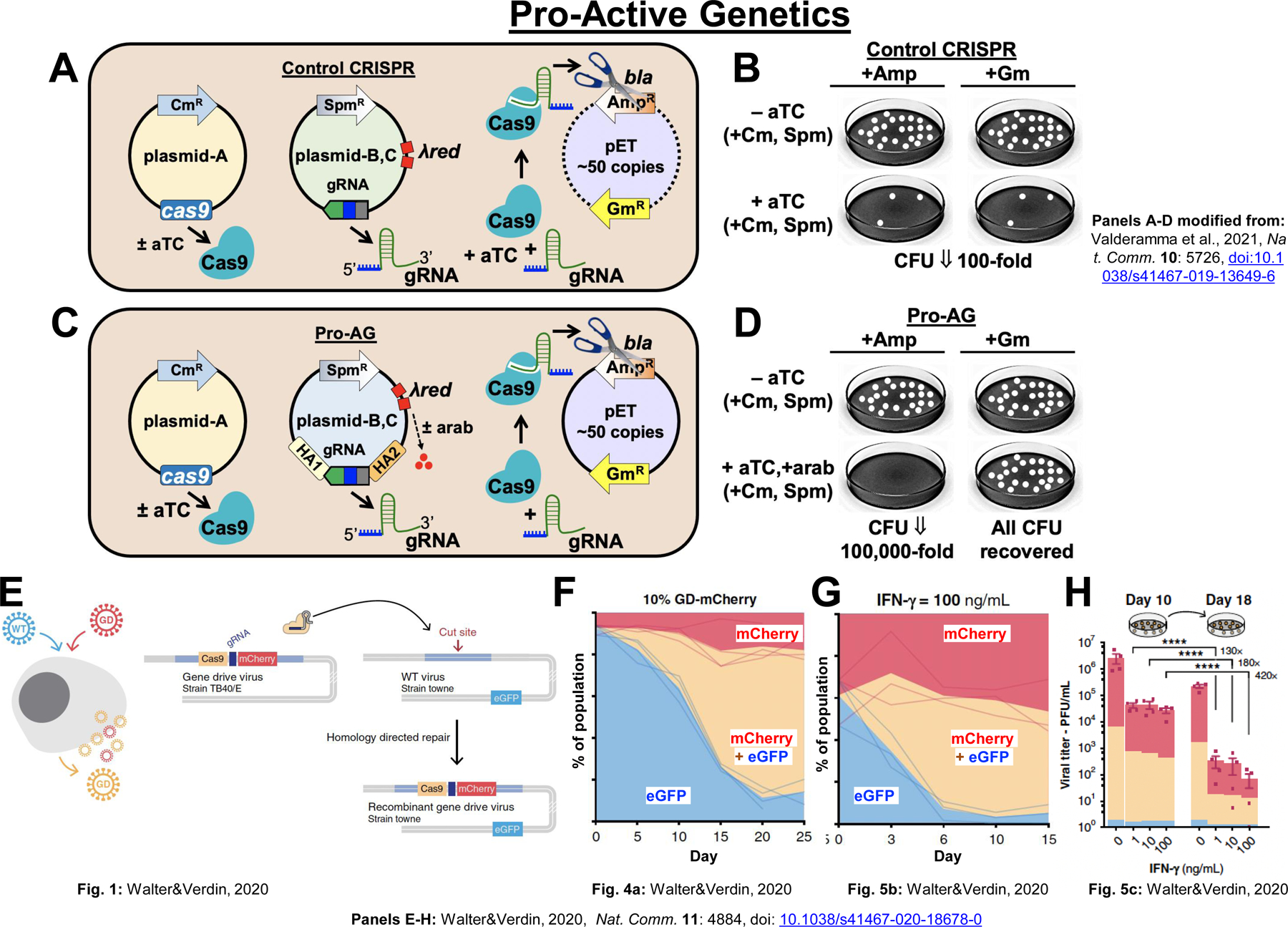
A) Diagram of control anti-AR CRISPR system components. A low copy-number plasmid-A (CmR) expresses Cas9 upon induction with anhydrotetracycline (aTC) and a low copy-number plasmid-BC (SpmR) expresses a gRNA constitutively (gray promoter box). A high-copy-number plasmid pET (AmpR, GmR) carries the AR target, beta lactamase (bla = AmpR), for gRNA-directed Cas9 cleavage. B) Control CRISPR CFU recovered on either Amp or Gm plates upon Cas9 induction are reduced by ~100-fold. C) Diagram of control Pro-AG system components. The pET (AmpR, GmR) target plasmid and Cas9-expressing plasmid are as in panel A. Plasmid B,C carries the gRNA flanked by beta lactamase homology arms (HA1 and HA2) and also carries a λred DNA repair cassette (red boxes), inducible by arabinose (arab, red dots). D) Induction of the Pro-AG system (+aTC, +arab) reduced CFU recovered by ~100,000-fold on Amp plates. Edited colonies could be quantitively recovered on Gm plates, however. E) Viral drive scheme. A gene-drive (GD) virus (human cytomegalovirus: hCMV) carries a cassette including Cas9, a gRNA specific, for cutting sequences in a wild-type (WT) target viral strain (hCMV-Towne) and a mCherry marker and is flanked by homology sequences specific for the target strain. The WT virus is marked with eGFP. Coinfection of cells with the GD and WT viruses results in copying of the drive cassette into the wild-type viral genome. F) Coinfection with the mCherry marked GD virus (10%) and eGFP marked WT target virus (90%) results in progressive copying of the drive cassette into the WT genome (double positive for mCherry and eGFP) to achieve ~90–95% final casette introgression. G) Treatment of human fibroblast cells with IFN-γ, which places the GF viral genome at a strong competitive disadvantage relative to WT (due to insertional inactivation of the UL23 gene), does not significantly alter the gene-drive trajectory compared to controls (no IFN-γ) during first generation viral growth due to the ability of WT virus to trans-complement the UL23 deficiency of the GD virus. H) During second generation (Day 18) infection under IFN-γ treatment without sufficient WT virus present to complement the GD virus, replication of the GD virus in greatly suppressed compared to first generation infections (Day 10).
Figure 5: Active Genetics in somatic cells.
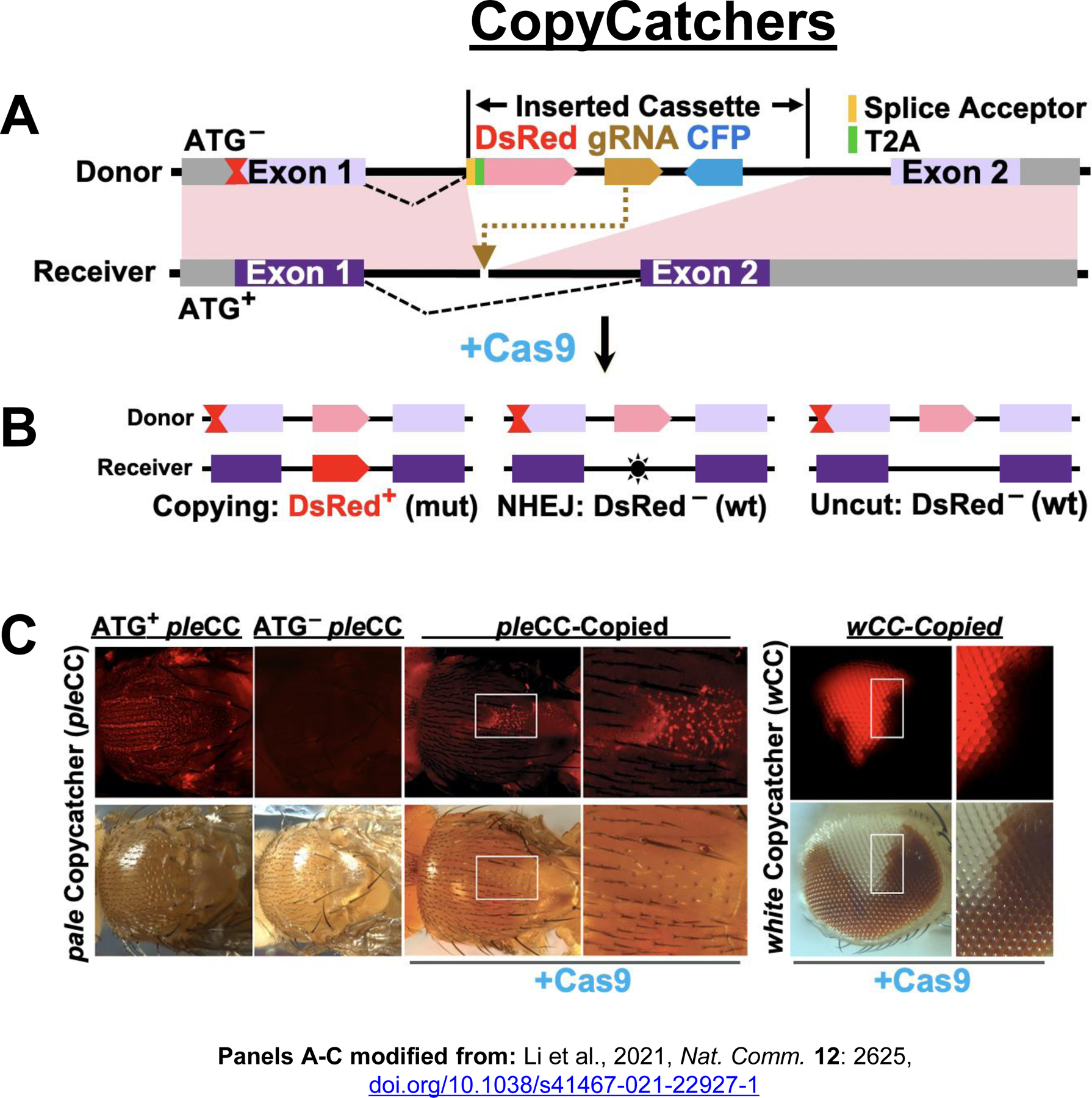
A) Scheme depicting a CopyCatcher element inserted into the intron of a target gene downstream of an associated ATG– mutation. CopyCatchers also carry gRNAs for copying themselves and a dominant CFP-eye marker. B) When combined with a separate source of Cas9 and a wild-type (wt) allele of the target gene, three outcomes are possible: copying (left), which separates the element from the ATG– mutation allowing for DsRed expression and generating a homozygous mutant condition; production of an NHEJ mutation (middle), or uncut alleles (right), the latter two resulting in unaltered phenotypes. C) Examples of CopyCatchers in the pale (ple) and white (w) loci. Left: When the ple, CopyCatcher (pleCC) is carried in a wild-type (ATG+) background, DsRed is expressed in nearly all cells giving rise to thoracic bristles (pleCC, wt) as a consequence of the T2A-Ds-Red cassette being spliced in-frame to the endogenous ple locus. When associated with an ATG– allele (pleCC:ATG–), however, DsRed expression is lost. DsRed expression can then be restored if the CopyCatcher element copies from the ATG– allele onto a homolog chromosome carrying a wild-type ATG+ allele (pleCC-copied). Tissue derived from such DsRed+ cells also display a homozygous mutant ple– phenotype owing to the function of both copies of the gene being disrupted. Right: representative w– clones reveal DsRed+ copying events in the eye for the white CopyCatcher (wCC).
Figure 6: Looking Forward.
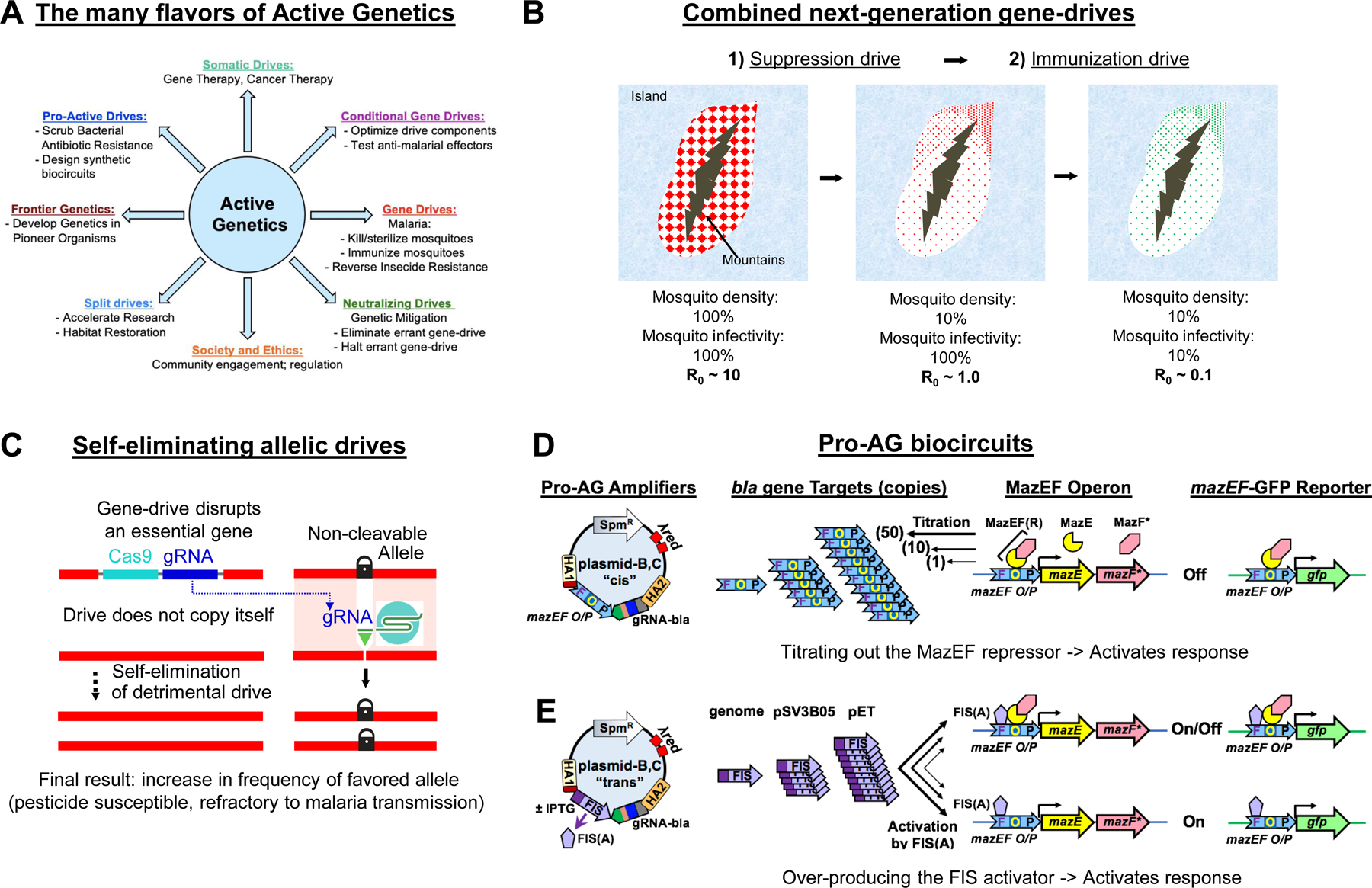
A) An updated wheel of potential active genetics applications pointing to new promising directions for this rapidly burgeoning field. Many of these avenues are extensions of lines of inquiry proposed in the previous 2016 review by Gantz and Bier in this same journal (see Fig. 1A). B) Sequential implementation of next-generation suppressive and modification (immunizing) gene-drives could cooperatively reduce the transmission of malaria. In the hypothetical example illustrated, the suppressive gene drive first reduces the number of malaria vectoring mosquitoes by ~90% (a value consistent with parameters derived from laboratory cage experiments [30,31] and modeling of potential target sites in Africa [133,134]) thereby reducing the average reproductive number (R0) for the parasite from an arbitrary initial value set at 10 for purposes of illustration (R0 values have been estimated to range from 1 – 3,000 [135]) to R0 ~1, below which malaria elimination is predicted under certain circumstances. Application of an efficient immunizing gene drive carrying effector molecules preventing transmission of malarial parasites [29,32] should then reduce the fraction of the remaining mosquitoes that are competent to serve as malarial vectors (in this case conservatively estimated again as a 90% reduction) leading to R0 falling well below the value required for efficient local elimination of malaria (e.g., R0 ~0.1). C) Self-eliminating allelic-drives could be devised in principle that are inserted into genes essential for viability or fertility that carry Cas9 and a gRNA targeting an unfavored allele located at another genomic site (e.g., the DTT and pyrethroid resistant 1014F allele of the voltage gated sodium ion channel), but not the preferred allele (e.g., the wild-type insecticide susceptible 1014L allele). Following introduction of insects carrying such a transiently acting drive element, the relative frequency of the favored allele would increase and then over time the drive element would disappear due to its heavy fitness cost. The result would be a non-transgenic (i.e., non-GMO) population of mosquitoes in which the relative frequency of a naturally occurring favored allele increased at the expense of an undesired allele. This soft touch process could be repeated as needed to maintain or increase the level of the favored allele. D,E) Proposed Pro-AG amplifier systems: cis (D) and trans (E). D) Donor plasmid-B,C “cis” carries the mazEF 77 bp operator/promoter as cargo in the <mazEF O/P gRNA-bla> cassette. By copying this cassette into low (genome = 1 copy), medium (pSV3B05 = 10–15 copies), or high (pET = ~50 copies) copy number targets differing levels of titration of the MazEF repressor should result in a threshold-dependent activation of both the endogenous MazEF operon and the MazEF-GFP reporter gene (inserted either into the genome or a plasmid (e.g., pET or pSV3B05) to vary its gene dosage. E) Donor plasmid-B,C “cis” carries a FIS transgene under control of the IPTG inducible lac promoter. FIS protein binds to site F in the mazEF promoter to activate transcription. Differing levels of FIS should then result in threshold dependent activation of both the endogenous MazEF operon and the MazEF-GFP reporter.
Gene-drives, allelic-drives and drive neutralizing elements
Gene-drives (Fig. 2A) and associated elements (Figs. 2B-E) represent one of the most impactful applications of active genetics. Since this topic has been recently reviewed in depth [26], only key findings will be summarized briefly here.
Next-generation gene-drive systems perform efficiently
The first kh modification drive element developed in mosquitoes mentioned above [16] and a prototype suppression drive developed in the laboratory of Andre Crisanti [27] were not efficiently transmitted via females due to Cas9/gRNA complexes that accumulated in the egg (reviewed in [26]). These maternally produced Cas9/gRNA complexes cleaved and mutated incoming paternal alleles following fertilization via the error-prone non-homologous end-joining (NHEJ) pathway. Multi-generational cage experiments conducted with the modification strain revealed that it actually behaved as a suppression drive. This kh-drive attained a high transient frequency in the population, but then often would lead to population collapse due to females being less viable and sterile if they inherited two mutant copies of the kh target gene [28]. Three potential solutions were envisioned to circumvent this challenge: 1) error-free drives that did not deposit appreciable levels of Cas9/gRNA complexes into the egg; 2) drive systems that killed or sterilized offspring inheriting mutant non-drive alleles (i.e., generated by the NHEJ pathway); or 3) male-only drives.
Second-generation gene-drive systems have been developed that overcome the female transmission problem via the first two imagined strategies (reviewed in more detail in [26]). With respect to the error-free approach, a modification-drive inserted into the cardinal locus in An. gambiae exhibits nearly optimal drive through both males and females using a particular promoter sequence from the germline determining gene nanos to express the Cas9 endonuclease [29]. The alternative strategy of eliminating copying errors that produce functional protected alleles of the target locus and impede spread of the drive has also culminated in highly efficient suppression drives. For example, the Crisanti group engineered sophisticated suppression drives inserted into in the doublesex locus of An. gambiae [30,31] that only produce non-functional NHEJ mutations, which also render females sterile. Similarly, a modification-drive carrying a recoded function-restoring allele of the kh locus drove efficiently by dominantly eliminating female progeny carrying non-functional NHEJ alleles via a process referred to as lethal mosaicism [32]. Lethal/sterile mosaicism relies on the gene-drive providing a functional recoded (non-cleavable) version of an essential target gene locus that is required for viability or fertility [18]. If such a drive element fails to be transmitted from mothers to their progeny, maternal Cas9/gRNA ribonucleoprotein complexes mutate the wild-type paternal allele in a substantial fraction of cells. Such mosaic individuals, which are rendered effectively homozygous mutant, are either in inviable or sterile depending on the required function of the essential gene. Analysis of gene-drives inserted into a variety of essential loci has validated the generality of this effective approach [32–34].
Two component-drives provide greater control and flexible deployment
An advantage of the bipartite nature of the Cas9/gRNA endonuclease is that it is possible to split the two core genetic entities (Cas9 and gRNA) between different elements (Fig. 2B). Such systems can either promote limited linear increases in the prevalence of the gRNA element of gRNA-only split drives endowed with the ability to copy in the presence of a Mendelian Cas9 source [33,35–38], or can be designed such that both components carry gRNAs to copy themselves, but only exhibit drive when combined, such as trans-complementing drives [39] (Fig. 2C) or integral drive systems developed by the groups of Windbichler and Christophides [40]. Each of these split elements alone, however, only exhibits “static” Mendelian inheritance.
The durability of split gRNA-only drives in a population can be tuned by adjusting initial release frequencies or the ratio of a Mendelian source of Cas9 to that of the drivable gRNA-only cassette [38]. Also, for some loci individuals carrying both the Cas9 and a gRNA element incur a transient drive-related fitness cost that results in disappearance of otherwise fitness neutral sources of Cas9 [38].
Split, dual driving elements offer the advantage of being able to test the activity of one or the other element under non-driving conditions (e.g., a cassette carrying anti-malarial effector proteins), which requires less stringent physical confinement than for full drives. Optimally performing components can then be assembled into a full-drive configuration.
Allelic drives increase prevalence of favored allelic variants
There are many polymorphic loci for which one allele is greatly preferred over another. In the case of insect pests, for example, genes encoding targets of insecticides are under selection to acquire mutations that render individuals insensitive to the action of such pesticides (typically by no longer binding the compound). Such resistance alleles have repeatedly emerged from treated populations [41]. Also, a naturally occurring viable allelic variant of the essential FREP1 locus in anopheline mosquitoes has been identified, which unlike the more common reference allele, fails to serve as a receptor to aid malarial parasites transit the intestinal epithelium into the body cavity, and thus does not sustain efficient parasite transmission [42–45].
Two types of strategies have been developed to increase transmission frequencies of preferred allelic variants of essential genes [18]. In the first, copy-cutting, a drive cassette (full or split) carries a second gRNA that selectively cleaves the unfavored allele (e.g., insecticide resistant or parasite enabling) (Fig. 2D). This simple approach results in the unfavored allele being cleaved and repaired using the preferred allele as a template. A limitation of this strategy is that it may not always be possible to design a gRNA with the required specificity for cleaving the unfavored allele. In a second more generally applicable approach, referred to as copy-grafting, is, one engineers a site resistant to gRNA cleavage in close proximity (within 25 bp) of the favored allelic variant. Drives carrying the gRNA cleave all alleles in general populations (favored or not), but will not cut the engineered cleavage resistant allele. Following cleavage of a sensitive undesired allele, and short range resection by the HDR repair pathway, localized gene-conversion events copy sequences including the adjacent preferred allele onto the homolog target chromosome. Non-functional NHEJ alleles that also may be generated are dominantly eliminated by lethal/sterile mosaicism. Copy-cutting and copy-grafting can both act with efficiencies comparable to those of gene-drives to bias inheritance of the preferred allele [18].
Proof-of principle has been provided in fruit flies for an allelic drive that reverses resistance to insecticides conferred by the 1014F allele of the voltage gated sodium ion channel [17]. An initial prevalence of 83% for the DDT resistant 1014F allele and 17% for the wild-type insecticide-susceptible 1014L allele was reversed to 13% 1014F and 87% wild-type 1014L after 10 generations as the driving cassette increased from 25% to ~80% prevalence in the population (e.g., as in Fig. 1D)..
Drive neutralizing elements efficiently replace or halt gene-drives
As summarized above, highly efficient second-generation gene-drive systems have now been developed that offer the potential to spread traits throughout insect populations. Concerns have been raised, however, regarding inherent potential risks associated with such highly invasive genetic elements. In our previous review, we proposed several strategies that have now been tested for eliminating or halting the spread of potentially undesired gene-drives.
One remediation approach, embodied in ERACRs (elements reversing the autocatalytic chain reaction) is to insert a gene cassette at the same site as a gene drive carrying two gRNAs that direct Cas9 cleavage on either side of the drive, but no Cas9 transgene of its own [1] (Fig. 2E, left panel). When an ERACR element, which cannot copy by itself, confronts a gene-drive element, the Cas9 provided by the drive cassette combines in trans with the gRNAs of ERACR to delete the drive element and copy the ERACR cassette in its place [19]. ERACRs also can carry cargo such as anti-malarial effectors or a recoded cDNA restoring activity of a locus whose function has been disrupted by insertion of the gene drive. In-depth analysis of one set of ERACRs in Drosophila revealed that these elements do not always perform as designed. For example, ERACRs can delete the drive without copying - often inducing chromosome damage, or alternatively, mutate the gRNA cut sites flanking the drive element leaving an active drive element intact and immune to further attack [19]. ERACRs damage target chromosomes because DNA sequence homology is lacking on one side of either gRNA cleavage site (i.e., the sides facing internal sequences of the gene cassette). Such a configuration is reminiscent of an aberrant Ds element studied McClintock that generated localized mutagenic events. These break-induced outcomes were similar to those produced by the process of chromothripsis [46,47], a mutagenic phenomenon that scrambles of the gene order of chromosomes and amplifies certain sequences in cancer cells, contributing further to tumorigenesis. Despite these imperfect DNA repair outcomes, ERACRs can efficiently delete and replace a gene-drive in population cages, particularly when given an added fitness advantage [19].
Another drive-neutralizing approach is to halt a drive by mutating and inactivating the Cas9 transgene that powers copying of the gene cassette. Two such drive-freezing systems have been tested: e-CHACRs (erasing constructs hitchhiking on the autocatalytic chain reaction) and Cas9-triggered chain ablation (CATCHA) elements [20]. e-CHACRs are gRNA-only cassettes that can be inserted anywhere in the genome. These elements carry a gRNA to copy themselves in the presence of Cas9 as well as one or more gRNAs to target Cas9 (Fig. 2E, right panels). Several e-CHACRs, inserted at different chromosomal locations have been evaluated, one of which efficiently eliminated a gene-drive in caged populations [19]. CATCHA elements, like an ERACR, are inserted at the same genomic location as a gene-drive, but similar to an e-CHACR, carry a gRNA targeting the Cas9 transgene leading to the CATCHA cassette inserting itself into the drive and inactivating it. Another strategy for inhibiting Cas9 activity developed by the Crisanti group, which is reversible, is to express Cas9-inhibitory proteins [21]. If a gene-drive carries a fitness cost, as suppression systems are designed to incur, it is also possible to replace the drive passively with a functional cleavage-resistant allele that imparts a selective advantage [28,48]. Finally, a strategy has been proposed by Adelman and colleagues to render Cas9-bearing gene-drive elements “biodegradable” by flanking them with repeated sequences and then encoding a second conditionally activatable nuclease that can cleave within the drive element to promote recombination between the repeats with consequent deletion of the internal drive cassette [49].
Looking Forward
Given the remarkable progress in developing second-generation gene-drives for population modification or suppression, the obvious next step is to test these systems under more realistic field conditions. One intriguing possibility is to use the two complementary drive approaches, suppression and modification, in tandem to reduce separate key parameters of disease transmission (vector density and infectivity, respectively) (Fig. 6B). Such tests could be conducted in large outdoor enclosed cages, which would retain much of the artificiality of laboratory cage experiments, or in a more relevant fashion in bounded geographic locations such as islands. It will also be important to develop detailed target product profiles (TPPs) that quantify key performance parameters such as drive efficiency, rate of off-target mutations, stability of the effector gene cassettes, and estimates of fitness costs specific for each of these various drives systems [29,50]. Most importantly, it is essential to continue current efforts to engage local, regional, and national stakeholders in open and transparent discussions of potential risks versus benefits of these next-generation drive systems or remediation strategies and what role these technologies might play in global efforts to eradicate vector borne diseases such as malaria [51,52].
On the technical front several challenges remain. One important objective will be to implement relevant technologies including allelic-drives and drive-neutralizing elements in disease vectors of key concern such as anopheline mosquitoes. Also, applications of more limited drive systems such as inherently self-limiting systems could offer effective tools to control both disease vectors and other insect pests on more local geographical scales. For example, one could insert an allelic-drive cassette into an essential gene so as disrupt its function and design the element to carry only a single gRNA that sustains allelic drive of a desired naturally occurring allele at a separate locus (e.g., insecticide susceptibility) (Fig. 6C). Release of such a fitness encumbered element should create a transient burst in the frequency of the preferred allele followed by the gene cassette being progressively eliminated from the population due to its fitness load. The net result would be only an alteration in the relative frequencies of two naturally occurring alleles, one of the very lightest touches possible for a gene-drive system.
Another technical challenge is to devise larger active genetic constructs that could accelerate genome engineering such as replacing several genome segments from one organism with those from another [53]. Currently, ~20 kb gene-drive elements have been observed to propagate with high efficiency [16]. Is there an upper limit to the size of copied cassettes? In organisms with many repeated genetic elements (such as transposons or repetitive coding or non-coding sequences) would such partial homology stretches lead to high rates of undesired recombination between such sequences as was observed at low frequency with ERACR elements that shared homology with their gene-drive targets [19]? Also, would it be possible to zip together a series of active genetic elements with overlapping sequences to create large contiguous genome insertions or replacements to permit facile modular assembly of multiple replaced loci? A recent full locus replacement of the Drosophila proboscipedia locus successfully employed such a strategy in a stepwise fashion [54]. Might it be possible also to accomplish such engineered tiling in a single step?
Generalizing active genetics to new organisms
While the great majority of progress in the gene-drive field has exploited established insect models (e.g., fruit flies and anopheline mosquitoes), efforts have also been initiated in other organisms including mosquitoes of the Culex genus [55] transmitting hemorrhagic viruses such as West Nile virus and Aedes genus [37] transmitting arboviruses such as Dengue, Chikungunya, and Zika. Moving deeper along the phylogenetic tree, prototype active genetic systems have also been developed in mammals (mice) [23,24], bacteria [25], and viruses [56] as summarized briefly below.
Mammalian drives underscore importance of limiting Cas9 activity to the germline
A prototype split-drive system similar to those implemented in insects has been developed in mice that results in significant super-Mendelian inheritance when transmitted through females (72% average transmission), but not via males [24]. Although the gender through which this drive copies efficiently is opposite to that observed in insects, the rationale for this difference is similar. In both classes of organisms the drive system performs best in the sex with fewest somatic cell cycles intervening between fertilization of the egg and segregation of the germline in the embryo (males in insects and females in mammals). Consistent with this hypothesis, delaying expression of Cas9 in male mice using a tightly controlled germline-expressed Cas9 source (under control of the spo11 promoter), resulted in similar, albeit modest, copying efficiencies through both sexes. The reduced overall copying in these experiments was due in part to the fact that most target alleles remained intact (i.e., wild-type) [23].
Bacterial drives can re-sensitize bacteria to antibiotics
Although CRISPR systems are prokaryotic in origin, most bacteria are not inherently equipped to sustain efficient HDR-based gene-drive type systems due to their being relatively poor in repairing double-strand breaks (DSBs) [25,57–59]. Indeed, the fraction of colony forming units (CFU) recovered following Cas9/gRNA induced DSBs in the bacterial genome is on the order of 10–8 [25]. Despite the dramatic difference in efficiencies of basal rates of DSB repair in bacteria versus eukaryotes, a conditional drive-like system referred to as Pro-Active Genetics (Pro-AG) has been developed in bacteria in which a gRNA bearing gene cassette flanked by homology arms can efficiently copy into homologous gene targets in the presence of DNA repair enzymes encoded by the phage-derived λ-Red system [25]. When targeting a beta-lactamase gene conferring resistance to the antibiotic ampicillin carried on a high copy plasmid, CRISPR strategies that cut and destroy plasmid targets can reduce antibiotic resistant (AR) CFU by ~100–1,000 fold (Fig. 3A,B) [25,57,58,60]. In contrast, the Pro-AG system decreased AR by a factor of ~105 (Fig. 3C,D) [25], greater than a hundred-fold improvement over the control CRISPR system. An important mechanism underlying self-amplification of the Pro-AG system is a positive feedback cycle based on increasing levels of the gRNA component as the Pro-AG element copies from a low copy-number donor plasmid into its high-copy number recipient target [25].
Viral gene-drives can reduce viral infectivity
Another promising avenue for CRISPR-based drives in mammalian cells is a gene-drive system developed to copy gene cassettes between two human cytomegalovirus (herpesvirus family) members [56]. In this system, developed by Marius Walter and Eric Verdin, a drive cassette inserted into the genome of one virus carries transgenes encoding Cas9, a gRNA designed to cut a second viral genome, and a fluorescent marker, which are flanked by homology arms adjacent to the gRNA target site in the second virus (Fig. 3E). Co-infection of cells with the two viruses leads to spread of the gene cassette from the donor virus to ~90–95% of the target viral genome and can result in replacement of the target genome in cultures cell populations (Fig. 3F). This system also can be used to reduce viral infectivity by targeting insertion sequences required conditionally (± IFN-γ treatment) for viral replication (Fig. 3G,H) offering potential new therapeutic avenues for treating and possibly curing persistent infections from this burdensome and sometimes lethal class of chronically infective viruses.
Looking Forward
Gene-drive systems similar to those used in mosquitoes may help combat a range of vector-borne diseases transmitted by other insects such as kissing bugs of the Rhodius genus which vector the pathogen causing Chagas disease [61], Tsetse flies, which carry trypanosome parasites responsible for sleeping sickness [62], and sand flies which vector Leishmaniasis [63]. Confinable gene-drive systems could also be used to combat diverse crop pests in well-delimited areas (e.g., specific cultivated fields).
Improving the efficiency of mammalian drive systems through both sexes could offer an important new avenue for restoring ecologies to a more native state (e.g., by eliminating invasive species for island conservation purposes [64–66]) or for preserving endangered species by immunizing them against threatening pathogens (e.g., expressing scFvs to prevent infection by bubonic plague in black-footed ferrets [67]). Analyzing drive efficiency using different gRNAs, varying levels and cell-type specificity of Cas9 expression, and genetic cassettes inserted at different loci and in different mammalian species (e.g., rats) should provide essential information for improving drive performance in these organisms. Conditional active genetic systems could also facilitate aggregation of complex traits in laboratory models (e.g. combining humanized gene modules in mice such as those used for optogenetic imaging or neuronal control). Similarly, generalization of such systems to plants could enable assembly of combinations of multiple desired alloalleles in polyploid crops, a task that remains extremely challenging using standard Mendelian approaches.
CRISPR-related approaches could prove invaluable in combatting the growing global concern of increasing antibiotic resistance in bacteria [68]. CRISPR-based systems summarized above offer potential advantages in this urgent effort as part of a rapidly diversifying toolbox of molecular components to develop ever more effective strategies to perform complex combinations of genome, episome, and phage edits to reduce the impact of antibiotic resistant pathogens and to promote homeostasis of healthy microbiomes. Among the key goals of such endeavors would be to scrub antibiotic resistance from both environmental and clinical settings, which could be augmented as mentioned above by harnessing horizontal gene transfer systems to disseminate anti-AR systems within proscribed bacterial populations.
Other types of bacterial elements might be combined with Pro-AG systems to increase their efficiency and extend their utility. For example, recently identified retrons are elements that carry bacterial reverse transcriptases, a single-strand DNA binding protein (SDP), and synthesize multicopy single-strand DNA (ssDNA) homology templates from non-coding RNA templates. These elements might be employed for parallel homology-based editing of multiple gene targets including genes encoding pathogenic virulence factors without the need to induce DSBs [69,70]. Retrons also exhibit anti-phage activities [71,72] that could be exploited to increase the fraction of bacteria in populations carrying gene-editing systems (i.e., by having phage kill cells that do not carry the editing cassette).
RNA-guided transposon systems [73–77] offer yet another potential mechanism for spreading a gene-casette of interest in bacteria. These targeted transposons carry nuclease-deficient Cas-related Cascade proteins that do not cut DNA, but instead direct the CRISPR-related complex to bind specific target sites in a gRNA-dependent fashion to mediate site specific transposition. If such transposons carried several gRNAs targeting multiple different sites, they should spread fluidly between these sites, some of which could be located on genetic entities such as phage or conjugal transfer elements to mediate horizontal gene transfer.
Synthetic biology is another exciting nascent field [78] that could exploit the potential of bacterial gene-editing systems (Fig. 6D). For example, the Pro-AG system can be viewed as an autonomous tunable solid-state amplifier circuit element. Since current strategies for creating amplifying circuits typically employ more cumbersome and fragile multistep transcriptional cascades, Pro-AG amplifiers could improve reliability and stability of these components[79,80]. Thus, one could insert genes encoding threshold-dependent transcription factors or titratable cis-acting regulatory sequences such as well-studied toxin-antitoxin regulatory systems [81] to provide various types of binary circuit outputs based on differing copy number of the target plasmid (Fig. 6E,F). It should also be possible to construct conditional sequential logic circuit elements based on one gene edit creating the required target site for a second edit. Shuttling such systems between different target elements using RNA-guided transposons or retrons to expand the range of allelic editing should fuel new rounds of innovation in this rapidly developing arena.
Active genetics could also play an enabling role in establishing new model genetic systems in organisms with few or no existing genetic tools. In addition to increasing the frequency of germline transmission of particular desired traits, active genetic systems could also include gRNAs expressed in somatic tissues (see next section) acting dominantly to produce readily identifiable phenotypes. Given the Herculean efforts typically required to establish new model genetic systems, active genetics could play an important role in accelerating such future endeavors.
CRISPR-based tools can be used for in vivo gene editing in somatic cells
CRISPR-based technologies have sparked a revolution in genome editing of somatic cells and tissues from various mammalian organisms. Edits are typically performed first by directing Cas9-dependent cleavage of specific genomic targets. Resulting DNA breaks are then repaired using co-transfected single-stranded or double-stranded correction templates. Although this breakthrough technology has many potential applications for engineering desired genetic changes in cultured cells, the requirement for exogenous DNA templates presents a significant barrier to performing in vivo gene editing. A notable exception is the use of CRISPR to delete or mutagenize sequences that exert dominant effects (Fig. 4A). Such efforts have led to successful gene therapies for diseases including muscular dystrophy in dogs [82] (where a defective exon in the dystrophin gene is deleted) and in humans for beta-thalassemia and sickle cell disease, (where a healthy embryonic hemoglobin gene homolog is reactivated in adulthood by mutagenizing a repressive cis-acting regulatory sequence [83]), as well as Leber’s congenital amaurosis 10 [84] and transthyretin amyloidosis [85] (where dominantly acting disease alleles are inactivated). These methods might also be employed to treat other dominantly acting mutations causing neurological diseases, such as Huntingtins [86,87]. Several more general strategies described below also have been developed to broaden the range and types of achievable in vivo gene editing.
Figure 4: In vivo gene editing approaches.
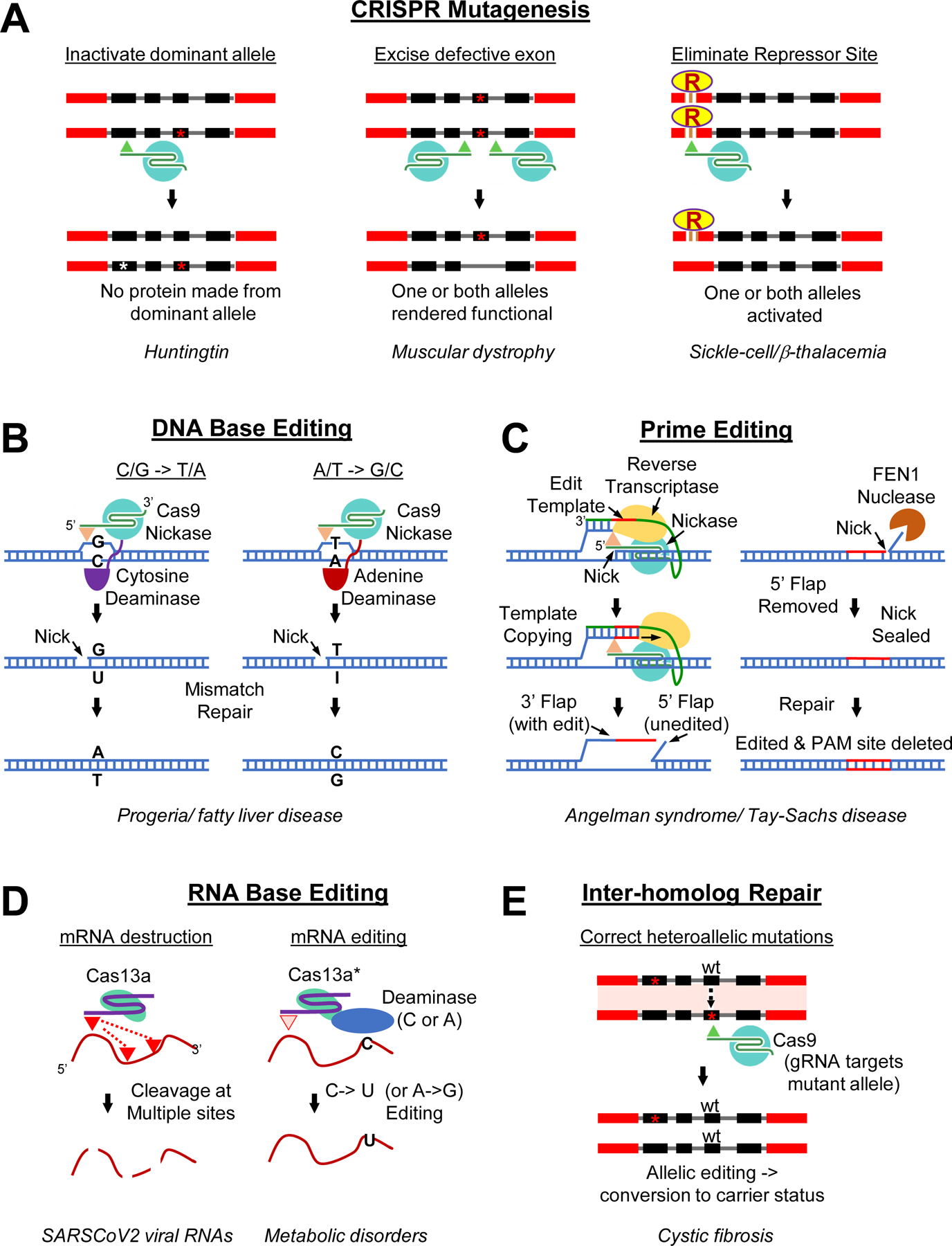
A) Supplying Cas9 and targeting gRNAs via viruses or other vectors can mutate and inactivate dominantly acting pathogenic alleles (left panel), delete (with two gRNAs) a defective non-essential exon (middle panel), or activate a quiescent therapeutically beneficial gene by eliminating a cis-acting repressor site that normally prevents the gene from being transcribed in the targeted cell type (right panel). Examples of diseases that either have been or could be treated by these strategies are indicated below the diagrams in italics for all panels of this figure. B) Scheme depicting concept of either cytosine (left panel) or adenine base editors. A mutant “Nickase” form of Cas9 nicks the DNA on the strand opposite to where the base alteration is to be made and is fused to deaminase enzymes that have been iteratively evolved to recognize DNA instead of RNA templates. These synthetic enzymes deaminate either cytosine or adenine residues within a narrow (~5 bp) window at the gRNA targeting site. The mismatch repair system can be biased to eliminate preferentially the residue on the nicked strand leading to its being replaced with the nucleotide complementary to the altered residue (either an A or a C) resulting in precise C/G -> T/A or A/T -> G/C edits. C) Scheme outlining the key features of prime editing. A Cas9 Nickase (Nickase) is fused to an RNA-directed reverse transcriptase and the gRNA is modified to carry a 3’ extension that can base pair with DNA sequences immediately 5’ to the nick to form a stable RNA/DNA hybrid primer. The extended gRNA also includes an editing template following the primer sequence that can then be copied into DNA by reverse transcriptase to create a 3’ Flap that can compete for hybridizing to the unaltered DNA strand (5’ Flap), displacing those sequences (at least transiently) where they can then be excised by the FEN1 nuclease. The resulting partially base-paired 3’ Flap can then be ligated to the 5’ end of the strand with the excised 5’ Flap and mismatch repair can result in editing that both incorporates the desired edit and also eliminates the endogenous PAM site (to prevent re-nicking). Addition of a separate Nickase that targets the non-altered strand (not shown in the diagram) at a short distance from the edited region can improve the efficiency of prime-editing via the same mechanism shown for base editing (i.e., biasing mismatch repair to favor using the non-nicked strand as a repair template). D) RNA-editing strategies include cleavage and destruction of specific mRNA templates (left panel) or editing the message (right panel). Destruction of targeted mRNAs is accomplished using RNA-directed nucleases such Cas13a which when complexed with a guide RNA are activated by binding to their specific targets. Once activated, the Cas13a nuclease cleaves the RNA in multiple locations and can also act in trans to cleave nearby RNA molecules (not depicted in the diagram). Editing of specific mRNAs can be achieved with enzymatically dead forms of Cas13a (Cas13a*) that have been fused to RNA directed deaminases (either cytosine or adenine deaminases) which result in editing specific residues (C-> U or A ->G). E) Inter-homolog repair of DSBs can take place in somatic cells either prior to DNA replication, when the homolog chromosome is the only available, or following DNA replication if the normal processes that favor repair from the identical sister chromosome are not operative (e.g., as caused by mutations resulting in loss of heterozygosity - LOH or by processes that favor pairing between chromosome homologs). In these scenarios, Cas9-induced DSBs can be repaired efficiently using the homolog as a template. The diagram illustrates a case in which two different loss-of-function alleles are present in a heteroallelic state. Allele specific cleavage of either mutant sequence can lead to repair from the non-cleaved functional sequence leading to restoration of endogenous gene activity controlled by native cis-regulatory elements on the repaired allele. Such repair if performed in a sufficient fraction of cells could convert an affected trans-heteroallelic patient into a carrier as depicted in the diagram or could repair a dominant-negatively acting allele (not shown).
DNA base-editing requires no added correction template
One approach to developing template free gene-editing systems suitable for in vivo applications has been to convert existing RNA-based editing enzymes into DNA-editors by iteratively evolving their specificities from RNA to DNA [88,89]. Such DNA editors, originally developed by Alexis Komor and colleagues in David Liu’s group, are fused to a cleavage-deficient form of Cas9 (enzymatically dead) to guide their mutagenic activities to specific genomic sites and changing residues to desired nucleotides within short editing windows of ~5 bp [90]. These tour de force efforts have resulted in so-called DNA base-editors that create specific types of edits (e.g., cytosine -> thymidine or adenine -> guanine) permitting correction of ~50% of possible desired mutant alleles (Fig. 4B).
Prime editing provides a platform with broad applications
A yet more general form of localized sequence editing also developed in the Liu lab uses a reverse transcriptase-Cas9 fusion and an altered gRNA (pegRNA) that includes a 5’ target site extension along with the desired templated sequence change [91]. In this system, referred to as prime editing (Fig. 4C), a Cas9 variant retains the ability to cleave one strand of the target to generate a nick at the editing site. The reverse transcriptase then primes synthesis of a single DNA strand using 5’-sequences of the pegRNA as a primer. As a result, the desired sequence change, which is incorporated into one strand, often converts the other strand by host cell DNA-repair pathways, a process that can be enhanced by further nicking of the opposite strand.
Base-editing strategies have also been extended to mitochondria by fusing the editing machinery to a protein domain that directs transport of the editor into mitochondria [92]. This breakthrough broadens therapeutic perspectives to the vast sphere of mitochondrial diseases and age-related disorders.
These stunning achievements in protein engineering and enzymatic repurposing have opened doors for new in vivo gene editing. For example, in the case of a pre-mature aging syndrome progeria, treatment of a mouse model for this disease [93] (these mice also age prematurely) resulted in a sufficient fraction of cells acquiring the desired corrective edit to endow those mice with longer healthy lives. Similarly, base editing inactivating the PCSK9 gene in the livers of mice and macaque monkeys resulted in the stable reduction of low-density lipoprotein (LDL) [94,95] (accomplished by upregulating levels of the LDL-receptor, which negatively regulates circulating LDL levels and is itself under negative control by PCSK9). Similar results in reducing LDL levels were also achieved using standard CRISPR-knockout of the PCSK9 gene [96].
RNA base-editing leaves the genome unaltered
A concern regarding CRISPR-based DNA editing is the possibility of generating off-target mutations that might exert long-lasting negative effects on patients. One approach to limit such potential risk is to employ target-specific RNA nucleases such as Cas13a either to degrade mRNA targets, or when fused to RNA-base editors [97], to edit specific RNAs (Fig. 4D). Such RNA-specific nucleases or modifying enzymes act in a transient fashion to reduce or alter target transcripts, respectively [98]. In the case of the native nucleases such as Cas13a, other random RNAs in close proximity to the nuclease also can be cleaved in trans once Cas13a has been activated by a gRNA binding to its target. In some cases, such pleiotropic targeting can lead to cell death. Treatment of retinal diseases by such downstream genetic interventions is one promising application of this technology [99].
RNA base editing may also prove useful in treating viral infections. For example, delivery of Cas13a and multiple gRNAs targeting the influenza and SARS-CoV2 genomes via a nebulizer reduced viral load significantly in mice [100].
Inter-homolog repair offers promise for hetero-allelic gene correction
When engaging the HDR pathway, somatic cells typically maintain genome integrity by employing identical sister chromatids as templates for DSB repair following DNA replication during the S and G2 phases of the cell cycle. This post-replicative and restorative function of HDR in somatic cells contrasts with the germline role of HDR where meiotic factors promote DSB-dependent recombination between homologous chromosomes [101–103]. In somatic cells, engagement of the homologous chromosome rather than the sister chromatid as the repair template (Fig. 4E) can be hazardous, resulting in Loss-of-Heterozygosity (LOH) phenotypes that can lead to oncogenic outcomes and developmental defects [104,105].
Although homologous chromosomes are not the favored DNA repair substrate in mammalian somatic cells [106], CRISPR-based editing experiments suggest that they may also serve as repair templates with varying degrees of efficiency [107]. There has been controversy over when during development such inter-homolog repair is likely to take place. A seminal study published by the Mitalipov group in 2017 [108] reported inter-homolog repair in early embryos possibly at the one or two cell stage. These findings, however, were questioned by several research groups who argued against the likelihood of such early inter-homolog repair due to homologous chromosomes being in separate cellular compartments [109–111]. Nonetheless, follow-up studies by the original group [112] provided additional support for their initial conclusions. Also, there is evidence for transient developmental windows during which alignment of homologs can take place [111], which correlate with periods during which significant frequencies of inter-homolog repair have been reported [113]. In this regard, it is relevant that DSBs have been observed to induce cytoskeletal-mediated re-localization of chromosomes within the nucleus [114,115] leading to regions of ~300 kb being aligned between homologous chromosomes following such DNA damage [116,117].
Recently, genetic sensors referred to as a CopyCatchers have been devised in Drosophila that permit detection of copying between chromosome homologs in somatic cells, while at the same time creating visible Loss-of-Function (LOF) mutant clones of the targeted locus [118] (Fig. 5A-C). These elements, which carry a gRNA targeting their own site of insertion, are inserted into introns of target genes and harbor a splice acceptor site that places a fluorescent marker in-frame with the endogenous locus, while simultaneously truncating the endogenous gene product (Fig. 5A). CopyCatchers are then combined with upstream LOF point-mutant alleles (e.g., ATG– mutations) that prevent expression of the fluorescent fusion protein. When these compound ATG– CopyCatcher alleles are placed in trans to a wild-type allele of the locus in the presence of a Cas9 source, the element can copy to the homologous chromosome (Fig. 5B), which then restores expression of the fluorescent marker (by separating it from the ATG– mutation) and at the same time creates clones of descendent cells with homozygous LOF phenotypes (Fig. 5C).
CopyCatchers inserted into two different pigmentation loci revealed surprisingly efficient inter-homolog copying (30%−50%) in cells of the epidermis and eye. Genetic modifier screens identified several loci which when knocked-down or over-expressed could alter copying rates [118]. One such modifier locus, which increased copying when knocked down, encodes the ortholog of the mammalian cMyc gene. In mammalian HEK293T cells, inter-homolog copying of an eGFP bearing gene cassette was also rigorously established, albeit at lower frequencies (4–8%) than observed in Drosophila. Importantly, genetic modifiers such as c-Myc that altered somatic cell gene conversion efficiencies in fruit flies also did so in mammalian cells[118].
Because mammalian chromosomes do not typically engage in inter-homolog pairing as pervasively as in somatic cells of Drosophila, there may be challenges in generalizing efficient inter-homolog copying to other organisms in which chromosome pairing is less prominent [119–121]. There is evidence in mammalian cells, however, that the absence of chromosome pairing is due in part to an active anti-pairing process, suggesting that these cells too might be induced to engage in pairing by interfering with this suppressive process [122]. Indeed, in some cancer cell lines, specific chromosome arms have been found to be consistently paired along their length [116,117,122]. Also, as mentioned above, the frequency of repairing DSB by copying from the homologous chromosome is greatly elevated in cells with mutations causing LOH [104,105], and in normal cells, DSBs result in transient localized paring of homologs [116,117]. Thus, with additional iterative advances, it may become possible to increase the efficiency of interhomolog repair in human somatic cells to revert disease causing alleles in patients carrying either a dominantly acting pathogenic allele or different heterozygous recessive LOF alleles. Such interventions could potentially convert affected patients into asymptomatic carriers (see more below in Looking Forward section).
Looking Forward
Perhaps the most salient promise of the various gene-editing approaches summarized above is in the arena of gene therapy wherein specific deleterious mutant alleles are reverted back to their original or a benign state in a sufficient proportion of somatic cells to reduce or possibly eliminate disease symptoms. As summarized above, a bevy of recent studies suggest that such therapeutics may become possible for treating several conditions, particularly when only a subset of cells need to be corrected to ameliorate symptoms. Base-editing or prime-editing systems offer the possibility of reverting specific mutations whether heterozygous or homozygous, while active genetic methods relying on inter-homolog repair would be applicable for patients carrying hetero-allelic LOF alleles. It might also be possible to combine these complementary approaches to increase the percentage of cells restored to a native state.
Another potential application of active genetic approaches is in the treatment of cancers in which amplified segments of the genome including over-expressed oncogenes are generated by the pathological process of chromothripsis [123–128]. Since chromothriptic chromosomes are reordered composites of chromosomal segments fused together [129], they carry novel sequences at those fusion sites which could in principle be targeted selectively by gRNAs that would not cut endogenous genome sequences.
Because overlapping but distinct genetic pathways mediate homology directed repair of DSBs during meiosis versus mitosis in somatic cells [101,102,130–132], it will be informative to employ the quantitative tools discussed above to identify new components through genetic screens that are either shared or unique to one or the other of these repair pathways. It may also be informative to exploit model systems to study DNA damage leading to chromothripsis. For example, as mentioned above, classic experiments of McClintock in maize lead to sectored pigmentation phenotypes in kernels. These mosaic phenotypes resulted from DSBs being induced by an active Ds element imbedded within second partial Ds element that thus lacked adjacent homology on the opposing homolog. Genetic screens for modifiers of such phenotypes could serve as a basis for identifying factors required for repair of these events, which may also overlap with factors involved in stitching back together chromosomes shattered by chromothripsis. Similarly, in-depth analysis of genes involved in creating or resolving the DNA damage induced by ERACR elements in fruit flies also may provide insights. These genetic studies could be complemented by targeted RNAi or CRISPRi knock-down of candidate genes in somatic cells encoding nucleases, ligases, and single-strand binding proteins that may play key roles in this fascinating phenomenon.
Conclusions
The promise of active genetics at its dawn in 2016 has been fulfilled in nearly all spheres initially envisioned including the development of efficient next-generation gene-drives, allelic-drives, drive neutralizing elements, and adaptation of these systems to other organisms including additional insect species, mammals, bacteria, and viruses. Similar copying of genetic elements to homologous chromosomes in somatic cells in combination with great advances in template-free genome editing such as base-editing and prime-editing also offer promise in the realm of next-generation gene therapies. Given the remarkable progress in these interdigitated arenas, the next five years are likely to witness a broad diversity of impactful applications for these now validated core methods, firmly establishing active genetics in the post-Mendelian era.
Box 1: Glossary.
Active Genetics:
Copying of a genetic element from one chromosome to its homologue in response to a double-strand DNA break being generated in the homologue at the same genomic site that the active genetic element in inserted. Copying, which results from directional gene conversion, is typically mediated in the germline by the synthesis-dependent strand annealing (SDSA or D-loop) branch of the homology-directed repair (HDR) pathway.
Aedes mosquitoes:
A genus of mosquitoes carrying arboviruses that transmit dengue fever, yellow fever, chikungunya, or Zika.
Allele:
Variant form of a gene that differs from an agreed upon reference DNA sequence.
Allelic-drives:
Genetic systems biasing the inheritance of a particular allelic variant, typically altering only one or a few base pairs. CRISPR-based allelic drive systems can be of two types, copy-cutting or copy-grafting.
Allo-alleles:
Alleles in a polyploid species (allopolyploid or autoploid) whose chromosomes either derive from whole-genome duplications within a species (autoploidy) or result from fusion of two different genomes (alloploidy) followed by potential partial loss of genetic information. Many crop species such as grains (wheat, rye), corn, peanuts, and sugarcane are polyploid as are a few animal species (e.g., Xenopus laevis African clawed frogs).
Anopheline mosquitoes:
A genus of mosquitoes that carry malarial parasites.
Anti-malarial effectors:
Transgenes encoding factors, which when expressed, exert a desired anti-malarial effect, such as for anti-malarial peptides expressed following a blood meal in mosquitoes.
Basic reproductive number (R0):
The expected number of disease cases generated by one person infected with a pathogen if all individuals are susceptible to infection (i.e., at the beginning of disease spread). For vector borne diseases, it depends on several factors involving the vector hosts as well as infected persons.
CHACRs:
(Constructs Hitchhiking on the Autocatalytic Chain Reaction). Active genetic elements carrying a guide RNA to copy themselves and potential cargo, but no Cas9 source.
Chromosomal translocations:
Rearrangements of the genome resulting from chromosomal segments breaking off from their original chromosomal locations and being rejoined to other chromosomal sites on the same or different chromosomes. In the case of reciprocal translocations, which can be viable and have normal fitness, breaks on two different chromosomal arms result in portions of one chromosome arm being switched for another to create two new chromosomes with the same gene complement present in the original strain.
Conjugal transfer:
Genetic entities that can be transmitted actively between bacteria. Examples include F-factors mediating transfer of plasmids or portions of the genome (high frequency recombination or HFR strains), which is a bacterial form of sexual reproduction. Full autonomous conjugal elements typically carry cis-acting OriT sequences required for their transfer to other cells as well as type IV secretory systems that encode the transfer machinery including pili through which the DNA is transferred. Many plasmids carry only OriT sequences and can only be transferred to recipient
CopyCatcher:
An active genetic element designed to detect homology directed copying from one chromosome to its homolog in somatic cells. CopyCatchers generate clonal fluorescent and mutant phenotypes when they copy.
Copy-cutting:
A form of allelic-drive in which the driving guide RNA directly cuts the non-preferred allelic variant leaving the preferred allele intact.
Copy-grafting:
A form of allelic-drive in which the preferred allele is adjacent to a sequence immune to cleavage by a guide RNA (gRNA) that can cut wild-type or non-preferred alleles. When the gRNA cuts either the wild-type or non-preferred allelic variants, the double strand break is repaired by homology-directed repair (HDR) using the cleavage-resistant allele as a template, copying the favored allelic variant in the process. The cleavage-resistant gRNA site and preferred allele are typically <25 bp apart to be copied reliably together as a biproduct of the short-range end-resection step of HDR.
CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats):
A bacterial immunity system from which a two component synthetic CRISPR/Cas9 genome editing system was derived.
Culex mosquitoes:
A genus of mosquitoes serving as vectors for diseases including: West Nile fever, St. Louis encephalitis, Japanese encephalitis, and viral diseases in birds and horses.
e-CHACR:
(Erasing Construct Hitchhiking on the Autocatalytic Chain Reaction). A drive-neutralizing active genetic element that encodes two or more guide RNAs (gRNAs) but no source of Cas9. One of the gRNAs cuts at the genomic site of e-CHACR insertion, enabling self-copying in the presence of a trans-acting source of Cas9 provided by the gene-drive. The additional gRNA(s) target cleavage and inactivation of the Cas9 transgene component of a gene-drive element. e-CHACRs can be inserted into the genome at any desired location.
ERACRs:
(Element Reversing the Autocatalytic Chain Reaction). Drive-neutralizing active genetic elements that encode two guide RNAs (gRNAs) cutting on either side of the gene-drive element. ERACRs are inserted at the same genomic site as the drive elements they are designed to delete and replace. ERACRs can carry additional cargo such as recoded copies of a gene targeted for disruption by the gene-drive. ERACRs do not carry a Cas9 source, however.
Essential gene:
A gene required for the viability or reproduction of an organism in a specific environment.
Gene-drive:
An allele of a diploid gene experiences drive if it is inherited more than 50% of the time (i.e., more than by random chance).
Guide RNA:
Expressed RNAs that bind to Cas9 and direct its cleavage of specific DNA targets. Endogenous bacterially expressed CRISPR guide RNAs consist of separate cRNAs and tracrRNAs that bind to Cas9 independently. Synthetic guide RNAs fuse the cRNAs and tracrRNAs into a single transcript, referred to as either a sgRNAs or more simply as gRNAs, the latter being the term used in this review.
Homology directed repair (HDR):
DNA repair pathway initiated following induction of double-strand DNA breaks in which the break is repaired by copying sequences (or a template) from an identical sister chromosome (typically following DNA replication in somatic cells) or from the homolog chromosome (typically during meiosis, although sometimes also in somatic cells).
HGT:
Horizontal gene transfer between individual organisms. In bacteria HGT can be mediated by naked DNA (transformation), conjugal transfer, transduction (by phage or phage-related entities including gene transfer agents), or by vesicular fusion.
Integral gene-drive:
Integral drives are comprised of two genetic components encoding the Cas9 and gRNA + effector inserted just upstream of the promoters of two different genes (loci 1 and 2). The Cas9 and effector genes each contain and intron into which the gRNA for copying that element is inserted and are followed by 2A ribosome-skipping sequence that leads to expression of Cas9 + endogenous locus-1 and the effector + endogenous locus-2. These two separate elements, like those of a trans-complementing drive are inherited in a Mendelian fashion, but copy together when combined thereby driving inheritance of each element.
Introduction:
The full introduction of a specific allele from one genetic background into another results in all progeny carrying the allele in question. The term introgression, typically of a trait that is potentially associated with additional surrounding local genetic variation from one strain into another background, can also be used in such contexts. Example 1: a gene-drive element carried in one strain that spreads to 100% introduction throughout a naïve target population. Example 2: an insecticide-resistant allele and a neighboring 1 megabase region were introgressed into sensitive strain of mosquitoes from a resistant strain.
Malaria:
An infectious blood disease caused by Plasmodial parasites (e.g., Plasmodium falciparum and Plasmodium vivax) that is primarily transmitted (vectored) by mosquitoes in the Anopheline genus.
Modification-drives:
Drive systems that modify a target population but do not reduce its numbers. Also referred to as replacement-drives.
Neutralizing-drives:
Genetic elements designed to eliminate or halt the spread of a gene-drive. Examples include active guide RNA (gRNA)-only elements such as Erasing Constructs Hitchhiking on the Autocatalytic Chain Reaction (e-CHACRs), Elements Reversing the Autocatalytic Chain Reaction (ERACRs), or Cas9-triggered chain ablation (CATCHA) elements, anti-Cas9 proteins, as well as passive recoded fitness-neutral gRNA target sites (for replacing suppression drives).
Non-homologous end-joining:
(NHEJ). A DNA repair pathway initiated following the induction of double-strand DNA breaks in which the two ends of DNA are ligated back to each other in a template-independent fashion. NHEJ can introduce small insertions or deletions, which can cause mutations and frameshift-based loss-of-function alleles of a gene.
Recoded-drives:
Gene-drives that carry a recoded version of a target gene into which it inserts such that the recoded sequences at the 5ʹ-end of the drive element are fused seamlessly to endogenous coding sequences, restoring wild-type protein activity and expression of the endogenous gene.
Risk assessment:
A formal analysis of all factors that need to be considered, evaluated and balanced before implementing an intervention such as a release of organisms carrying a gene-drive element. Risks should include predicted or potential consequences associated with performing the intervention as well as those accompanying a decision not to implement the technology.
Selfish genes:
Genes that are transmitted from parents to progeny in a biased fashion. See Super-Mendelian elements. Typically, selfish genes are naturally occurring entities.
Super-Mendelian elements:
Genetic entities in diploid eukaryotic organisms, such as gene-drives, selfish genes, or other active genetic elements, that are inherited more often than by chance alone (i.e., by > 50% of their offspring).
Suppression-drives:
Gene-drives that reduce the size of the target population. Typically, suppression drives insert into genes required for the viability or fertility of one or both sexes.
Trans-complementing drive:
A split CRISPR-based gene-drive system comprising two components: a Cas9-expressing element and a guide RNA (gRNA)-bearing element that are inserted at different sites in the genome. The gRNA element carries two gRNAs, one to copy itself and the other cutting at the insertion site of the Cas9 element. Alone, each element is inherited in a standard Mendelian fashion. When combined, however, the gRNAs complex with Cas9 provided in trans to drive copying of both elements thereby creating a full-drive system that efficiently transmits both elements to progeny.
Transformation:
Uptake of naked extracellular DNA sequences into bacterial cells and insertion of these sequences into the genome or episomal elements.
Abbreviations
- CATCHA
Cas9-triggered chain ablation
- CFU
colony forming unit
- CRISPR
Clustered Regularly Interspaced Short Palindromic Repeats
- DSB
double-strand break
- e-CHACR
erasing construct hitchhiking on the autocatalytic chain reaction
- ERACR
element reversing the autocatalytic chain reaction
- HDR
homology directed repair
- HEG
homing endonuclease gene
- gRNA
guide RNA
- LDL
low-density lipoprotein
- LOF
Loss-of-Function
- LOH
Loss of heterozygosity
- NHEJ
non-homologous end-joining
- Pro-AG
Pro-Active Genetics
- TPP
Target Product Profile
Footnotes
Conflict of Interest: Ethan Bier and Valentino Gantz have equity interest in two companies they co-founded: Synbal Inc. and Agragene, Inc., which may potentially benefit from the research results summarized in this review. They also serve on Synbal’s board of directors and the scientific advisory board for both companies.
References
- [1].Gantz VM, & Bier E (2016). The dawn of active genetics. Bioessays, 38(1), 50–63. doi: 10.1002/bies.201500102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Gantz VM, & Bier E (2015). The mutagenic chain reaction: a method for converting heterozygous to homozygous mutations. Science, 348(6233), 442–444. doi: science.aaa5945 [pii] 10.1126/science.aaa5945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Comfort NC (2001). The Tangled Field: Barbara McClintock’s search for patterns of genetic controll Cambridge, Massachusetts: Harvard University Press. [Google Scholar]
- [4].Doring HP, & Starlinger P (1984). Barbara McClintock’s controlling elements: now at the DNA level. Cell, 39(2 Pt 1), 253–259. doi: 10.1016/0092-8674(84)90002-3 [DOI] [PubMed] [Google Scholar]
- [5].Fedoroff N, Wessler S, & Shure M (1983). Isolation of the transposable maize controlling elements Ac and Ds. Cell, 35(1), 235–242. doi: 10.1016/0092-8674(83)90226-x [DOI] [PubMed] [Google Scholar]
- [6].Chevalier BS, & Stoddard BL (2001). Homing endonucleases: structural and functional insight into the catalysts of intron/intein mobility. Nucleic Acids Res, 29(18), 3757–3774. doi: 10.1093/nar/29.18.3757 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Macreadie IG, Scott RM, Zinn AR, & Butow RA (1985). Transposition of an intron in yeast mitochondria requires a protein encoded by that intron. Cell, 41(2), 395–402. doi: 10.1016/s0092-8674(85)80012-x [DOI] [PubMed] [Google Scholar]
- [8].Kruger AN, & Mueller JL (2021). Mechanisms of meiotic drive in symmetric and asymmetric meiosis. Cell Mol Life Sci, 78(7), 3205–3218. doi: 10.1007/s00018-020-03735-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Lyon MF (2003). Transmission ratio distortion in mice. Annu Rev Genet, 37, 393–408. doi: 10.1146/annurev.genet.37.110801.143030 [DOI] [PubMed] [Google Scholar]
- [10].Sugimoto M (2014). Developmental genetics of the mouse t-complex. Genes Genet Syst, 89(3), 109–120. doi: 10.1266/ggs.89.109 [DOI] [PubMed] [Google Scholar]
- [11].Curtis CF (1968). Possible use of translocations to fix desirable genes in insect pest populations. Nature, 218(5139), 368–369. [DOI] [PubMed] [Google Scholar]
- [12].Burt A (2003). Site-specific selfish genes as tools for the control and genetic engineering of natural populations. Proc Biol Sci, 270(1518), 921–928. doi: 10.1098/rspb.2002.2319 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Chan YS, Huen DS, Glauert R, Whiteway E, & Russell S (2013). Optimising homing endonuclease gene drive performance in a semi-refractory species: the Drosophila melanogaster experience. PLoS One, 8(1), e54130. doi: 10.1371/journal.pone.0054130PONE-D-12-25967 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Rong YS, & Golic KG (2003). The homologous chromosome is an effective template for the repair of mitotic DNA double-strand breaks in Drosophila. Genetics, 165(4), 1831–1842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Windbichler N, Menichelli M, Papathanos PA, Thyme SB, Li H, Ulge UY, . . . Crisanti A (2011). A synthetic homing endonuclease-based gene drive system in the human malaria mosquito. Nature, 473(7346), 212–215. doi: nature09937 [pii] 10.1038/nature09937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Gantz VM, Jasinskiene N, Tatarenkova O, Fazekas A, Macias VM, Bier E, & James AA (2015). Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proc Natl Acad Sci U S A, 112(49), E6736–6743. doi: 1521077112 [pii] 10.1073/pnas.1521077112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Kaduskar B, Kushwah RBS, Auradkar A, Guichard A, Li M, Bennett JB, . . . Bier E (2022). Reversing insecticide resistance with allelic-drive in Drosophila melanogaster. Nat Commun, 13(1), 291. doi: 10.1038/s41467-021-27654-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Guichard A, Haque T, Bobik M, Xu XS, Klanseck C, Kushwah RBS, . . . Bier E (2019). Efficient allelic-drive in Drosophila. Nat Commun, 10(1), 1640. doi: 10.1038/s41467-019-09694-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Xu XS, Bulger EA, Gantz VM, Klanseck C, Heimler SR, Auradkar A, . . . Bier E (2020). Active genetic neutralizing elements for halting or deleting gene drives. Mol Cell, 80(2), 246–262. doi: 10.1016/j.molcel.2020.09.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Wu B, Luo L, & Gao XJ (2016). Cas9-triggered chain ablation of cas9 as a gene drive brake. Nat Biotechnol, 34(2), 137–138. doi: 10.1038/nbt.3444 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Taxiarchi C, Beaghton A, Don NI, Kyrou K, Gribble M, Shittu D, . . . Crisanti A (2021). A genetically encoded anti-CRISPR protein constrains gene drive spread and prevents population suppression. Nat Commun, 12(1), 3977. doi: 10.1038/s41467-021-24214-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Liu JZ, Ali SR, Bier E, & Nizet V (2018). Innate Immune Interactions between Bacillus anthracis and Host Neutrophils. Front Cell Infect Microbiol, 8, 2. doi: 10.3389/fcimb.2018.00002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Weitzel AJ, Grunwald HA, Weber C, Levina R, Gantz VM, Hedrick SM, . . . Cooper KL (2021). Meiotic Cas9 expression mediates gene conversion in the male and female mouse germline. PLoS Biol, 19(12), e3001478. doi: 10.1371/journal.pbio.3001478 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Grunwald HA, Gantz VM, Poplawski G, Xu XS, Bier E, & Cooper KL (2019). Super-Mendelian inheritance mediated by CRISPR-Cas9 in the female mouse germline. Nature, 566(7742), 105–109. doi: 10.1038/s41586-019-0875-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Valderrama JA, Kulkarni SS, Nizet V, & Bier E (2019). A bacterial gene-drive system efficiently edits and inactivates a high copy number antibiotic resistance locus. Nature Communications, 10(1), 5726. doi: 10.1038/s41467-019-13649-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Bier E (2022). Gene drives gaining speed. Nat Rev Genet, 23(1), 5–22. doi: 10.1038/s41576-021-00386-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Hammond A, Galizi R, Kyrou K, Simoni A, Siniscalchi C, Katsanos D, . . . Nolan T (2016). A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nat Biotechnol, 34(1), 78–83. doi: 10.1038/nbt.3439 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Pham TB, Phong CH, Bennett JB, Hwang K, Jasinskiene N, Parker K, . . . James AA (2019). Experimental population modification of the malaria vector mosquito, Anopheles stephensi. PLoS Genet, 15(12), e1008440. doi: 10.1371/journal.pgen.1008440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Carballar-Lejarazu R, Ogaugwu C, Tushar T, Kelsey A, Pham TB, Murphy J, . . . James AA (2020). Next-generation gene drive for population modification of the malaria vector mosquito, Anopheles gambiae. Proc Natl Acad Sci U S A doi: 10.1073/pnas.2010214117 [DOI] [PMC free article] [PubMed]
- [30].Simoni A, Hammond AM, Beaghton AK, Galizi R, Taxiarchi C, Kyrou K, . . . Crisanti A (2020). A male-biased sex-distorter gene drive for the human malaria vector Anopheles gambiae. Nat Biotechnol doi: 10.1038/s41587-020-0508-1 [DOI] [PMC free article] [PubMed]
- [31].Kyrou K, Hammond AM, Galizi R, Kranjc N, Burt A, Beaghton AK, . . . Crisanti A (2018). A CRISPR-Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat Biotechnol, 36(11), 1062–1066. doi: 10.1038/nbt.4245 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Adolfi A, Gantz VM, Jasinskiene N, Lee H-F, Hwang K, Bulger EA, . . . James AA (2020). Efficient population modification gene-drive rescue system in the malaria mosquito Anopheles stephensi. Nat Commun(11), 5553. doi: 10.1038/s41467-020-19426-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Kandul NP, Liu J, Bennett JB, Marshall JM, & Akbari OS (2021). A confinable home-and-rescue gene drive for population modification. Elife, 10. doi: 10.7554/eLife.65939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Terradas G, Buchman AB, Bennett JB, Shriner I, Marshall JM, Akbari OS, & Bier E (2021). Inherently confinable split-drive systems in Drosophila. Nature Communications, 12(1), 1480. doi: 10.1038/s41467-021-21771-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Kandul NP, Liu J, Buchman A, Gantz VM, Bier E, & Akbari OS (2020). Assessment of a Split Homing Based Gene Drive for Efficient Knockout of Multiple Genes. G3 (Bethesda), 10(2), 827–837. doi: 10.1534/g3.119.400985 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Champer J, Yang E, Lee E, Liu J, Clark AG, & Messer PW (2020). A CRISPR homing gene drive targeting a haplolethal gene removes resistance alleles and successfully spreads through a cage population. Proc Natl Acad Sci U S A, 117(39), 24377–24383. doi: 10.1073/pnas.2004373117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Li M, Yang T, Kandul NP, Bui M, Gamez S, Raban R, . . . Akbari OS (2020). Development of a confinable gene drive system in the human disease vector Aedes aegypti. Elife, 9. doi: 10.7554/eLife.51701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Terradas G, Buchman AB, Bennett JB, Shriner I, Marshall JM, Akbari OS, & Bier E (2021). Inherently confinable split-drive systems in Drosophila. Nat Commun, 12(1), 1480. doi: 10.1038/s41467-021-21771-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].López Del Amo V, Bishop AL, Sanchez CH, Bennett JB, Feng X, Marshall JM, . . . Gantz VM (2020). A transcomplementing gene drive provides a flexible platform for laboratory investigation and potential field deployment. Nat Commun, 11(1), 352. doi: 10.1038/s41467-019-13977-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Nash A, Urdaneta GM, Beaghton AK, Hoermann A, Papathanos PA, Christophides GK, & Windbichler N (2019). Integral gene drives for population replacement. Biol Open, 8(1). doi: 10.1242/bio.037762 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Clarkson CS, Miles A, Harding NJ, O’Reilly AO, Weetman D, Kwiatkowski D, . . . Anopheles gambiae Genomes, C. (2021). The genetic architecture of target-site resistance to pyrethroid insecticides in the African malaria vectors Anopheles gambiae and Anopheles coluzzii. Mol Ecol doi: 10.1111/mec.15845 [DOI] [PMC free article] [PubMed]
- [42].Li J, Wang X, Zhang G, Githure JI, Yan G, & James AA (2013). Genome-block expression-assisted association studies discover malaria resistance genes in Anopheles gambiae. Proc Natl Acad Sci U S A, 110(51), 20675–20680. doi: 1321024110 [pii] 10.1073/pnas.1321024110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Niu G, Franc AC, Zhang G, Roobsoong W, Nguitragool W, Wang X, . . . Li J (2017). The fibrinogen-like domain of FREP1 protein is a broad-spectrum malaria transmission-blocking vaccine antigen. J Biol Chem, 292(28), 11960–11969. doi: 10.1074/jbc.M116.773564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Zhang G, Niu G, Franca CM, Dong Y, Wang X, Butler NS, . . . Li J (2015). Anopheles Midgut FREP1 Mediates Plasmodium Invasion. J Biol Chem, 290(27), 16490–16501. doi: 10.1074/jbc.M114.623165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Dong Y, Simoes ML, Marois E, & Dimopoulos G (2018). CRISPR/Cas9 -mediated gene knockout of Anopheles gambiae FREP1 suppresses malaria parasite infection. PLoS Pathog, 14(3), e1006898. doi: 10.1371/journal.ppat.1006898 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Shorokhova M, Nikolsky N, & Grinchuk T (2021). Chromothripsis-Explosion in Genetic Science. Cells, 10(5). doi: 10.3390/cells10051102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Krupina K, Goginashvili A, & Cleveland DW (2021). Causes and consequences of micronuclei. Curr Opin Cell Biol, 70, 91–99. doi: 10.1016/j.ceb.2021.01.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Hammond AM, Kyrou K, Bruttini M, North A, Galizi R, Karlsson X, . . . Nolan T (2017). The creation and selection of mutations resistant to a gene drive over multiple generations in the malaria mosquito. PLoS Genet, 13(10), e1007039. doi: 10.1371/journal.pgen.1007039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Zapletal J, Najmitabrizi N, Erraguntla M, Lawley MA, Myles KM, & Adelman ZN (2021). Making gene drive biodegradable. Philos Trans R Soc Lond B Biol Sci, 376(1818), 20190804. doi: 10.1098/rstb.2019.0804 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].James SL, Marshall JM, Christophides GK, Okumu FO, & Nolan T (2020). Toward the Definition of Efficacy and Safety Criteria for Advancing Gene Drive-Modified Mosquitoes to Field Testing. Vector Borne Zoonotic Dis, 20(4), 237–251. doi: 10.1089/vbz.2019.2606 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Kormos A, Lanzaro GC, Bier E, Dimopoulos G, Marshall JM, Pinto J, . . . James AA (2020). Application of the Relationship-Based Model to Engagement for Field Trials of Genetically Engineered Malaria Vectors. Am J Trop Med Hyg doi: 10.4269/ajtmh.20-0868 [DOI] [PMC free article] [PubMed]
- [52].James S, Collins FH, Welkhoff PA, Emerson C, Godfray HCJ, Gottlieb M, . . . Toure YT (2018). Pathway to Deployment of Gene Drive Mosquitoes as a Potential Biocontrol Tool for Elimination of Malaria in Sub-Saharan Africa: Recommendations of a Scientific Working Group(dagger). Am J Trop Med Hyg, 98(6_Suppl), 1–49. doi: 10.4269/ajtmh.18-0083 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Xu XS, Gantz VM, Siomava N, & Bier E (2017). CRISPR/Cas9 and active genetics-based trans-species replacement of the endogenous Drosophila kni-L2 CRM reveals unexpected complexity. Elife, 6. doi: 10.7554/eLife.30281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Auradkar A, Bulger EA, Devkota S, McGinnis W, & Bier E (2021). Dissecting the evolutionary role of the Hox gene proboscipedia in Drosophila mouthpart diversification by full locus replacement. Sci Adv, 7(46), eabk1003. doi: 10.1126/sciadv.abk1003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Feng X, Lopez Del Amo V, Mameli E, Lee M, Bishop AL, Perrimon N, & Gantz VM (2021). Optimized CRISPR tools and site-directed transgenesis towards gene drive development in Culex quinquefasciatus mosquitoes. Nat Commun, 12(1), 2960. doi: 10.1038/s41467-021-23239-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Walter M, & Verdin E (2020). Viral gene drive in herpesviruses. Nat Commun, 11(1), 4884. doi: 10.1038/s41467-020-18678-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Bikard D, Euler CW, Jiang W, Nussenzweig PM, Goldberg GW, Duportet X, . . . Marraffini LA (2014). Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials. Nat Biotechnol, 32(11), 1146–1150. doi: 10.1038/nbt.3043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Citorik RJ, Mimee M, & Lu TK (2014). Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases. Nat Biotechnol, 32(11), 1141–1145. doi: 10.1038/nbt.3011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Yosef I, Manor M, Kiro R, & Qimron U (2015). Temperate and lytic bacteriophages programmed to sensitize and kill antibiotic-resistant bacteria. Proc Natl Acad Sci U S A, 112(23), 7267–7272. doi: 10.1073/pnas.1500107112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Kim JS, Cho DH, Park M, Chung WJ, Shin D, Ko KS, & Kweon DH (2016). CRISPR/Cas9-Mediated Re-Sensitization of Antibiotic-Resistant Escherichia coli Harboring Extended-Spectrum beta-Lactamases. J Microbiol Biotechnol, 26(2), 394–401. doi: 10.4014/jmb.1508.08080 [DOI] [PubMed] [Google Scholar]
- [61].Lidani KCF, Andrade FA, Bavia L, Damasceno FS, Beltrame MH, Messias-Reason IJ, & Sandri TL (2019). Chagas Disease: From Discovery to a Worldwide Health Problem. Front Public Health, 7, 166. doi: 10.3389/fpubh.2019.00166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Buscher P, Cecchi G, Jamonneau V, & Priotto G (2017). Human African trypanosomiasis. Lancet, 390(10110), 2397–2409. doi: 10.1016/S0140-6736(17)31510-6 [DOI] [PubMed] [Google Scholar]
- [63].Desjeux P (2004). Leishmaniasis: current situation and new perspectives. Comp Immunol Microbiol Infect Dis, 27(5), 305–318. doi: 10.1016/j.cimid.2004.03.004 [DOI] [PubMed] [Google Scholar]
- [64].Teem JL, Alphey L, Descamps S, Edgington MP, Edwards O, Gemmell N, . . . Roberts A (2020). Genetic Biocontrol for Invasive Species. Front Bioeng Biotechnol, 8, 452. doi: 10.3389/fbioe.2020.00452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Godwin J, Serr M, Barnhill-Dilling SK, Blondel DV, Brown PR, Campbell K, . . . Thomas P (2019). Rodent gene drives for conservation: opportunities and data needs. Proc Biol Sci, 286(1914), 20191606. doi: 10.1098/rspb.2019.1606 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].McFarlane GR, Whitelaw CBA, & Lillico SG (2018). CRISPR-Based Gene Drives for Pest Control. Trends Biotechnol, 36(2), 130–133. doi: 10.1016/j.tibtech.2017.10.001 [DOI] [PubMed] [Google Scholar]
- [67].Salkeld DJ (2017). Vaccines for Conservation: Plague, Prairie Dogs & Black-Footed Ferrets as a Case Study. Ecohealth, 14(3), 432–437. doi: 10.1007/s10393-017-1273-6 [DOI] [PubMed] [Google Scholar]
- [68].Bier E, & Nizet V (2021). Driving to Safety: CRISPR-Based Genetic Approaches to Reducing Antibiotic Resistance. Trends Genet doi: 10.1016/j.tig.2021.02.007 [DOI] [PubMed]
- [69].Lim H, Jun S, Park M, Lim J, Jeong J, Lee JH, & Bang D (2020). Multiplex Generation, Tracking, and Functional Screening of Substitution Mutants Using a CRISPR/Retron System. ACS Synth Biol, 9(5), 1003–1009. doi: 10.1021/acssynbio.0c00002 [DOI] [PubMed] [Google Scholar]
- [70].Farzadfard F, & Lu TK (2014). Synthetic biology. Genomically encoded analog memory with precise in vivo DNA writing in living cell populations. Science, 346(6211), 1256272. doi: 10.1126/science.1256272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Millman A, Bernheim A, Stokar-Avihail A, Fedorenko T, Voichek M, Leavitt A, . . . Sorek R (2020). Bacterial Retrons Function In Anti-Phage Defense. Cell doi: 10.1016/j.cell.2020.09.065 [DOI] [PubMed]
- [72].Gao L, Altae-Tran H, Bohning F, Makarova KS, Segel M, Schmid-Burgk JL, . . . Zhang F (2020). Diverse enzymatic activities mediate antiviral immunity in prokaryotes. Science, 369(6507), 1077–1084. doi: 10.1126/science.aba0372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Klompe SE, Vo PLH, Halpin-Healy TS, & Sternberg SH (2019). Transposon-encoded CRISPR-Cas systems direct RNA-guided DNA integration. Nature, 571(7764), 219–225. doi: 10.1038/s41586-019-1323-z [DOI] [PubMed] [Google Scholar]
- [74].Koonin EV, Makarova KS, Wolf YI, & Krupovic M (2020). Evolutionary entanglement of mobile genetic elements and host defence systems: guns for hire. Nat Rev Genet, 21(2), 119–131. doi: 10.1038/s41576-019-0172-9 [DOI] [PubMed] [Google Scholar]
- [75].Strecker J, Ladha A, Gardner Z, Schmid-Burgk JL, Makarova KS, Koonin EV, & Zhang F (2019). RNA-guided DNA insertion with CRISPR-associated transposases. Science, 365(6448), 48–53. doi: 10.1126/science.aax9181 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Peters JE, Makarova KS, Shmakov S, & Koonin EV (2017). Recruitment of CRISPR-Cas systems by Tn7-like transposons. Proc Natl Acad Sci U S A, 114(35), E7358–E7366. doi: 10.1073/pnas.1709035114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Wiegand T, & Wiedenheft B (2020). CRISPR Surveillance Turns Transposon Taxi. CRISPR J, 3(1), 10–12. doi: 10.1089/crispr.2020.29081.twi [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Bashor CJ, & Collins JJ (2018). Understanding Biological Regulation Through Synthetic Biology. Annu Rev Biophys, 47, 399–423. doi: 10.1146/annurev-biophys-070816-033903 [DOI] [PubMed] [Google Scholar]
- [79].Bashor CJ, Patel N, Choubey S, Beyzavi A, Kondev J, Collins JJ, & Khalil AS (2019). Complex signal processing in synthetic gene circuits using cooperative regulatory assemblies. Science, 364(6440), 593–597. doi: 10.1126/science.aau8287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [80].Lee JW, Chan CTY, Slomovic S, & Collins JJ (2018). Next-generation biocontainment systems for engineered organisms. Nat Chem Biol, 14(6), 530–537. doi: 10.1038/s41589-018-0056-x [DOI] [PubMed] [Google Scholar]
- [81].Chan WT, Espinosa M, & Yeo CC (2016). Keeping the Wolves at Bay: Antitoxins of Prokaryotic Type II Toxin-Antitoxin Systems. Front Mol Biosci, 3, 9. doi: 10.3389/fmolb.2016.00009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Amoasii L, Hildyard JCW, Li H, Sanchez-Ortiz E, Mireault A, Caballero D, . . . Olson EN (2018). Gene editing restores dystrophin expression in a canine model of Duchenne muscular dystrophy. Science, 362(6410), 86–91. doi: 10.1126/science.aau1549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Frangoul H, Altshuler D, Cappellini MD, Chen YS, Domm J, Eustace BK, . . . Corbacioglu S (2021). CRISPR-Cas9 Gene Editing for Sickle Cell Disease and beta-Thalassemia. N Engl J Med, 384(3), 252–260. doi: 10.1056/NEJMoa2031054 [DOI] [PubMed] [Google Scholar]
- [84].Ledford H (2020). CRISPR treatment inserted directly into the body for first time. Nature, 579(7798), 185. doi: 10.1038/d41586-020-00655-8 [DOI] [PubMed] [Google Scholar]
- [85].Gillmore JD, Gane E, Taubel J, Kao J, Fontana M, Maitland ML, . . . Lebwohl D (2021). CRISPR-Cas9 In Vivo Gene Editing for Transthyretin Amyloidosis. N Engl J Med doi: 10.1056/NEJMoa2107454 [DOI] [PubMed]
- [86].Monteys AM, Ebanks SA, Keiser MS, & Davidson BL (2017). CRISPR/Cas9 Editing of the Mutant Huntingtin Allele In Vitro and In Vivo. Mol Ther, 25(1), 12–23. doi: 10.1016/j.ymthe.2016.11.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].McMahon MA, & Cleveland DW (2017). Gene therapy: Gene-editing therapy for neurological disease. Nat Rev Neurol, 13(1), 7–9. doi: 10.1038/nrneurol.2016.190 [DOI] [PubMed] [Google Scholar]
- [88].Anzalone AV, Koblan LW, & Liu DR (2020). Genome editing with CRISPR-Cas nucleases, base editors, transposases and prime editors. Nat Biotechnol, 38(7), 824–844. doi: 10.1038/s41587-020-0561-9 [DOI] [PubMed] [Google Scholar]
- [89].Rees HA, & Liu DR (2018). Base editing: precision chemistry on the genome and transcriptome of living cells. Nat Rev Genet, 19(12), 770–788. doi: 10.1038/s41576-018-0059-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Komor AC, Badran AH, & Liu DR (2018). Editing the Genome Without Double-Stranded DNA Breaks. ACS Chem Biol, 13(2), 383–388. doi: 10.1021/acschembio.7b00710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Anzalone AV, Randolph PB, Davis JR, Sousa AA, Koblan LW, Levy JM, . . . Liu DR (2019). Search-and-replace genome editing without double-strand breaks or donor DNA. Nature, 576(7785), 149–157. doi: 10.1038/s41586-019-1711-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Mok BY, de Moraes MH, Zeng J, Bosch DE, Kotrys AV, Raguram A, . . . Liu DR (2020). A bacterial cytidine deaminase toxin enables CRISPR-free mitochondrial base editing. Nature, 583(7817), 631–637. doi: 10.1038/s41586-020-2477-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [93].Koblan LW, Erdos MR, Gordon LB, Collins FS, Brown JD, & Liu DR (2021). Base editor treats progeria in mice. Nature doi: 10.1038/d41586-021-01114-8 [DOI] [PubMed]
- [94].Musunuru K, Chadwick AC, Mizoguchi T, Garcia SP, DeNizio JE, Reiss CW, . . . Kathiresan S (2021). In vivo CRISPR base editing of PCSK9 durably lowers cholesterol in primates. Nature, 593(7859), 429–434. doi: 10.1038/s41586-021-03534-y [DOI] [PubMed] [Google Scholar]
- [95].Rothgangl T, Dennis MK, Lin PJC, Oka R, Witzigmann D, Villiger L, . . . Schwank G (2021). In vivo adenine base editing of PCSK9 in macaques reduces LDL cholesterol levels. Nat Biotechnol doi: 10.1038/s41587-021-00933-4 [DOI] [PMC free article] [PubMed]
- [96].Wang L, Breton C, Warzecha CC, Bell P, Yan H, He Z, . . . Wilson JM (2021). Long-term stable reduction of low-density lipoprotein in nonhuman primates following in vivo genome editing of PCSK9. Mol Ther, 29(6), 2019–2029. doi: 10.1016/j.ymthe.2021.02.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [97].Zhang F (2019). Exploration of Microbial Diversity to Discover Novel Molecular Technologies. Keio J Med, 68(1), 26. doi: 10.2302/kjm.68-002-ABST [DOI] [PubMed] [Google Scholar]
- [98].Xu C, Zhou Y, Xiao Q, He B, Geng G, Wang Z, . . . Yang H (2021). Programmable RNA editing with compact CRISPR–Cas13 systems from uncultivated microbes. Nature Methods, 18(5), 499–506. doi: 10.1038/s41592-021-01124-4 [DOI] [PubMed] [Google Scholar]
- [99].Fry LE, Peddle CF, Barnard AR, McClements ME, & MacLaren RE (2020). RNA editing as a therapeutic approach for retinal gene therapy requiring long coding sequences. Int J Mol Sci, 21(3). doi: 10.3390/ijms21030777 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Blanchard EL, Vanover D, Bawage SS, Tiwari PM, Rotolo L, Beyersdorf J, . . . Santangelo PJ (2021). Treatment of influenza and SARS-CoV-2 infections via mRNA-encoded Cas13a in rodents. Nat Biotechnol, 39(6), 717–726. doi: 10.1038/s41587-021-00822-w [DOI] [PubMed] [Google Scholar]
- [101].Bier E, Harrison MM, O’Connor-Giles KM, & Wildonger J (2018). Advances in Engineering the Fly Genome with the CRISPR-Cas System. Genetics, 208(1), 1–18. doi: 10.1534/genetics.117.1113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Haber JE (2015). TOPping off meiosis. Mol Cell, 57(4), 577–581. doi: 10.1016/j.molcel.2015.02.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Kadyk LC, & Hartwell LH (1992). Sister chromatids are preferred over homologs as substrates for recombinational repair in Saccharomyces cerevisiae. Genetics, 132(2), 387–402. doi: 10.1093/genetics/132.2.387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [104].Ceccaldi R, Rondinelli B, & D’Andrea AD (2016). Repair Pathway Choices and Consequences at the Double-Strand Break. Trends Cell Biol, 26(1), 52–64. doi: 10.1016/j.tcb.2015.07.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [105].Prakash R, Zhang Y, Feng W, & Jasin M (2015). Homologous recombination and human health: the roles of BRCA1, BRCA2, and associated proteins. Cold Spring Harb Perspect Biol, 7(4), a016600. doi: 10.1101/cshperspect.a016600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [106].Stark JM, & Jasin M (2003). Extensive loss of heterozygosity is suppressed during homologous repair of chromosomal breaks. Mol Cell Biol, 23(2), 733–743. doi: 10.1128/MCB.23.2.733-743.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [107].Kosicki M, Tomberg K, & Bradley A (2018). Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements. Nat Biotechnol, 36(8), 765–771. doi: 10.1038/nbt.4192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [108].Ma H, Marti-Gutierrez N, Park SW, Wu J, Lee Y, Suzuki K, . . . Mitalipov S (2017). Correction of a pathogenic gene mutation in human embryos. Nature, 548(7668), 413–419. doi: 10.1038/nature23305 [DOI] [PubMed] [Google Scholar]
- [109].Egli D, Zuccaro MV, Kosicki M, Church GM, Bradley A, & Jasin M (2018). Inter-homologue repair in fertilized human eggs? Nature, 560(7717), E5–E7. doi: 10.1038/s41586-018-0379-5 [DOI] [PubMed] [Google Scholar]
- [110].Adikusuma F, Piltz S, Corbett MA, Turvey M, McColl SR, Helbig KJ, . . . Thomas PQ (2018). Large deletions induced by Cas9 cleavage. Nature, 560(7717), E8–E9. doi: 10.1038/s41586-018-0380-z [DOI] [PubMed] [Google Scholar]
- [111].Reichmann J, Nijmeijer B, Hossain MJ, Eguren M, Schneider I, Politi AZ, . . . Ellenberg J (2018). Dual-spindle formation in zygotes keeps parental genomes apart in early mammalian embryos. Science, 361(6398), 189–193. doi: 10.1126/science.aar7462 [DOI] [PubMed] [Google Scholar]
- [112].Ma H, Marti-Gutierrez N, Park SW, Wu J, Hayama T, Darby H, . . . Mitalipov S (2018). Ma et al. reply. Nature, 560(7717), E10–E23. doi: 10.1038/s41586-018-0381-y [DOI] [PubMed] [Google Scholar]
- [113].Wilde JJ, Aida T, Del Rosario RCH, Kaiser T, Qi P, Wienisch M, . . . Feng G (2021). Efficient embryonic homozygous gene conversion via RAD51-enhanced interhomolog repair. Cell, 184(12), 3267–3280 e3218. doi: 10.1016/j.cell.2021.04.035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [114].Caridi CP, D’Agostino C, Ryu T, Zapotoczny G, Delabaere L, Li X, . . . Chiolo I (2018). Nuclear F-actin and myosins drive relocalization of heterochromatic breaks. Nature, 559(7712), 54–60. doi: 10.1038/s41586-018-0242-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [115].Oshidari R, Strecker J, Chung DKC, Abraham KJ, Chan JNY, Damaren CJ, & Mekhail K (2018). Nuclear microtubule filaments mediate non-linear directional motion of chromatin and promote DNA repair. Nat Commun, 9(1), 2567. doi: 10.1038/s41467-018-05009-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [116].Gandhi M, Evdokimova VN, Cuenco KT, Bakkenist CJ, & Nikiforov YE (2013). Homologous chromosomes move and rapidly initiate contact at the sites of double-strand breaks in genes in G(0)-phase human cells. Cell Cycle, 12(4), 547–552. doi: 10.4161/cc.23754 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [117].Gandhi M, Evdokimova VN, K, T. C., Nikiforova MN, Kelly LM, Stringer JR, . . . Nikiforov YE (2012). Homologous chromosomes make contact at the sites of double-strand breaks in genes in somatic G0/G1-phase human cells. Proc Natl Acad Sci U S A, 109(24), 9454–9459. doi: 10.1073/pnas.1205759109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [118].Li Z, Marcel N, Devkota S, Auradkar A, Hedrick SM, Gantz VM, & Bier E (2021). CopyCatchers are versatile active genetic elements that detect and quantify inter-homolog somatic gene conversion. Nature Communications, 12(1), 2625. doi: 10.1038/s41467-021-22927-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [119].Joyce EF, Williams BR, Xie T, & Wu CT (2012). Identification of genes that promote or antagonize somatic homolog pairing using a high-throughput FISH-based screen. PLoS Genet, 8(5), e1002667. doi: 10.1371/journal.pgen.1002667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [120].Lewis E (1954). The theory and application of a new method of detecting chromosomal rearrangements in Drosophila melanogaster. Amer. Naturalist, 88(841), 225–239. [Google Scholar]
- [121].Metz CW (1916). Chromosome studies on the Diptera. II. The paired association of chromosomes in the Diptera, and its significance. J. Exp. Zool(21), 213–280. doi: 10.1002/jez.1400210204 [DOI] [Google Scholar]
- [122].Joyce EF, Erceg J, & Wu CT (2016). Pairing and anti-pairing: a balancing act in the diploid genome. Curr Opin Genet Dev, 37, 119–128. doi: 10.1016/j.gde.2016.03.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [123].Wu S, Bafna V, & Mischel PS (2021). Extrachromosomal DNA (ecDNA) in cancer pathogenesis. Curr Opin Genet Dev, 66, 78–82. doi: 10.1016/j.gde.2021.01.001 [DOI] [PubMed] [Google Scholar]
- [124].Zeng X, Wan M, & Wu J (2020). ecDNA within tumors: a new mechanism that drives tumor heterogeneity and drug resistance. Signal Transduct Target Ther, 5(1), 277. doi: 10.1038/s41392-020-00403-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [125].Kim H, Nguyen NP, Turner K, Wu S, Gujar AD, Luebeck J, . . . Verhaak RGW (2020). Extrachromosomal DNA is associated with oncogene amplification and poor outcome across multiple cancers. Nat Genet, 52(9), 891–897. doi: 10.1038/s41588-020-0678-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [126].Verhaak RGW, Bafna V, & Mischel PS (2019). Extrachromosomal oncogene amplification in tumour pathogenesis and evolution. Nat Rev Cancer, 19(5), 283–288. doi: 10.1038/s41568-019-0128-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [127].Turner KM, Deshpande V, Beyter D, Koga T, Rusert J, Lee C, . . . Mischel P (2017). Extrachromosomal oncogene amplification drives tumour evolution and genetic heterogeneity. Nature, 543(7643), 122–125. doi: 10.1038/nature21356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [128].Shoshani O, Brunner SF, Yaeger R, Ly P, Nechemia-Arbely Y, Kim DH, . . . Cleveland DW (2021). Chromothripsis drives the evolution of gene amplification in cancer. Nature, 591(7848), 137–141. doi: 10.1038/s41586-020-03064-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- [129].Wu S, Turner KM, Nguyen N, Raviram R, Erb M, Santini J, . . . Mischel PS (2019). Circular ecDNA promotes accessible chromatin and high oncogene expression. Nature, 575(7784), 699–703. doi: 10.1038/s41586-019-1763-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [130].Enguita-Marruedo A, Martin-Ruiz M, Garcia E, Gil-Fernandez A, Parra MT, Viera A, . . . Page J (2019). Transition from a meiotic to a somatic-like DNA damage response during the pachytene stage in mouse meiosis. PLoS Genet, 15(1), e1007439. doi: 10.1371/journal.pgen.1007439 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [131].Goedecke W, Eijpe M, Offenberg HH, van Aalderen M, & Heyting C (1999). Mre11 and Ku70 interact in somatic cells, but are differentially expressed in early meiosis. Nat Genet, 23(2), 194–198. doi: 10.1038/13821 [DOI] [PubMed] [Google Scholar]
- [132].McKee BD (2004). Homologous pairing and chromosome dynamics in meiosis and mitosis. Biochim Biophys Acta, 1677(1–3), 165–180. doi: 10.1016/j.bbaexp.2003.11.017 [DOI] [PubMed] [Google Scholar]
- [133].North AR, Burt A, & Godfray HCJ (2020). Modelling the suppression of a malaria vector using a CRISPR-Cas9 gene drive to reduce female fertility. BMC Biol, 18(1), 98. doi: 10.1186/s12915-020-00834-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- [134].North AR, Burt A, & Godfray HCJ (2019). Modelling the potential of genetic control of malaria mosquitoes at national scale. BMC Biol, 17(1), 26. doi: 10.1186/s12915-019-0645-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [135].Smith DL, McKenzie FE, Snow RW, & Hay SI (2007). Revisiting the basic reproductive number for malaria and its implications for malaria control. PLoS Biol, 5(3), e42. doi: 10.1371/journal.pbio.0050042 [DOI] [PMC free article] [PubMed] [Google Scholar]