

Abstract
Consumption and its excesses are sometimes explained by imbalance of need or lack of control over “wanting.” “Wanting” assigns value to cues that predict rewards, whereas “needing” assigns value to biologically significant stimuli that one is deprived of. Here we aimed at studying how the brain activation patterns related to value of “wanted” stimuli differs from that of “needed” stimuli using activation likelihood estimation neuroimaging meta‐analysis approaches. We used the perception of a cue predicting a reward for “wanting” related value and the perception of food stimuli in a hungry state as a model for “needing” related value. We carried out separate, contrasts, and conjunction meta‐analyses to identify differences and similarities between “wanting” and “needing” values. Our overall results for “wanting” related value show consistent activation of the ventral tegmental area, striatum, and pallidum, regions that both activate behavior and direct choice, while for “needing” related value, we found an overall consistent activation of the middle insula and to some extent the caudal‐ventral putamen, regions that only direct choice. Our study suggests that wanting has more control on consumption and behavioral activation.
Keywords: consumption, fMRI, motivation, needing, wanting

1. INTRODUCTION
Current consumption (e.g., food, transport, etc.) in western countries seems to be one of the causes of ecological problems we are facing (Lipschutz, 2001). According to some, this consumption is in part due to the fact that we consume what we want beyond what we need (Stern, 2000). Apart from problems related to ecology, in our daily behaviors such as those related to food consumption, excesses, and maladaptive behaviors are sometimes explained by an imbalance of need or a lack of control over “wanting.” Indeed, Campbell (1998) reports that research on consumer behavior has shown that there are two types of rhetoric used to justify the action of purchase: needs and wants, as well as their synonyms. He also argues that rhetoric of needs is derived from utilitarianism and puritanism that advocated a life based on necessity or satisfaction, while the rhetoric of wants is based on romanticism and linked to the pursuit of pleasure (Campbell, 1998). Beyond this rhetorical distinction, one can wonder: is there a true difference between something that we need and something that we want? At the neural level, needing is related to a state of deprivation of something important for life or survival (Bouton, 2016), and increases arousal through interoceptive salience (Craig, 2003). “Wanting” is related to the prediction of reward in the brain and is more closely related to motivation (Berridge, 2004). Although earlier theories suggested that need or deprivation defines motivation (Hull, 1943), it was later demonstrated that cues that signal hunger do not elicit motivation to eat, while food‐related cues did lead to motivation to eat (Bindra, 1974), showing that motivated behaviors are more determined by reward prediction (which is closer to “wanting”) than need state, though the latter can have a multiplicative effect (Toates, 1994). The distinction might seem clear. However, since “needed” stimuli are often pursued and thus associated with motivational value, it is not obvious whether an external stimulus, such as food, is pursued because of its “needing” or “wanting” related value. As “needed” stimuli might have a different form of value than “wanted” stimuli, we can wonder how in the brain the value of “needed” stimuli differs from that of “wanted” stimuli.
Given that we cannot test wanting and/or needing as general phenomena, here we are testing some manifestation of them. Hence, to conceptualize this distinction, we refer to “wanting a stimulus” as “Wanting”ST, which represents a brain reaction to a reward predicting cue that triggers reward seeking. We refer to “needing a stimulus” as “Needing”ST, which represents the brain reaction to a stimulus that one is deprived of, without necessarily seeking it. We propose to use activation pattern to the perception of a cue predicting a reward as a model for “Wanting”ST related value, and to use the activation pattern during the perception of a food stimulus while in the state of hunger as a model for “Needing”ST related value (Silverman et al., 2014; Spear, 2011). In order to answer our question, we used a neuroimaging meta‐analytic approach, comparing the patterns of brain activations during the processing of “Wanting”ST versus “Needing”ST. Previous meta‐analyses have focused on either “Wanting”ST (Oldham et al., 2018; Sescousse et al., 2013; Wilson et al., 2018) or “Needing”ST (Chen & Zeffiro, 2020; LaBar et al., 2001; van der Laan et al., 2011), but no work has directly compared both activation patterns.
1.1. “Wanting”ST related value
Nonhuman animals tend to respond and “want” food even when they are no longer hungry (Bouton, 2016). This is also the case for humans: cues of M&M or pictures of cigarettes (for smokers) lead to more consumption even after having been consumed to satiety (Hogarth & Chase, 2011; Watson et al., 2014). “Wanting”ST is a concept from incentive salience theory that comes from animal studies (Berridge & Robinson, 1998; see also: Ikemoto & Panksepp, 1999; Salamone & Correa, 2002; Salamone et al., 1997), which states that “Wanting”ST is based on two neuropsychological processes: the first is a pavlovian cue that predicts the reward, and the second is the dopaminergic state (which might be enhanced by hunger, thirst, emotions, drugs, etc.) (Berridge, 1996). In other words, “Wanting”ST depends on external stimuli that act as pavlovian cues that predict rewards (Berridge, 2018). The attribution of value to these cues depends on mesolimbic dopamine (Berridge, 1996). The latter is secreted within the ventral tegmental area (VTA) by a reward cue (Schultz, 1998) and projected within the nucleus accumbens (Nacc), ventral pallidum, and on the central amygdala opioid (Warlow & Berridge, 2021; Zhang et al., 2009). In this sense, the VTA would be more related to reward prediction (Schultz, 2015; Schultz et al., 1997) and its phasic activation determines directional value (preference/choice or action selection), while the NAcc is more related to value attribution (an incentive salience) to that reward prediction (Berridge & Aldridge, 2009; Hamid et al., 2016; Lex & Hauber, 2008), along with the central nucleus of the amygdala (Balleine & Killcross, 2006; Warlow & Berridge, 2021; Zhang et al., 2009). Thus, “Wanting”ST starts with reward prediction. Human studies on reward prediction have shown the involvements of the orbitofrontal cortex (OFC) (O'Doherty, 2004; O'Doherty et al., 2002), VTA (Carter et al., 2009; Krebs et al., 2009; O'Doherty et al., 2002; Oldham et al., 2018; Schott et al., 2008), NAcc and ventral striatum (Carter et al., 2009; Knutson et al., 2001, 2003; O'Doherty, 2004; O'Doherty et al., 2002; Oldham et al., 2018; Schott et al., 2008; Simon et al., 2015; Wilson et al., 2018), amygdala (O'Doherty, 2004; O'Doherty et al., 2002; Oldham et al., 2018), and insula (O'Doherty et al., 2002; Oldham et al., 2018; Wilson et al., 2018). These results suggest that, in humans, the activation pattern of reward prediction which leads to “Wanting”ST related value could implicate these regions.
1.2. “Needing”ST related value
A need state has the capacity to give and to control the preference/choice related value of a novel food or drink in relation to its consequence on the organism, once the organism has experienced the benefit of that stimulus in the need state (Balleine, 1992; Dickinson & Balleine, 1994). Thus, “Needing”ST related value can have an impact on choice and action selection (Dickinson & Balleine, 1994) or preference (Salamone et al., 2018). For instance, hunger influences flavor preference learning in humans based on flavor (Brunstrom & Fletcher, 2008; Zellner et al., 1983), nutrients (Appleton et al., 2006; Gibson et al., 1995; Kern et al., 1993), and odor‐sweetness (Yeomans & Mobini, 2006). Moreover, the shifts in preference are found to persist beyond the initial training period (Brunstrom & Fletcher, 2008), suggesting a long‐term learned value. However, though “Needing”ST provides directional value (preference/choice or action selection), in absence of reward prediction, “Needing”ST (by itself) does not activate behavior (Berridge, 2004; Bindra, 1974; Bolles & Moot, 1972; Toates, 1994). In the brain, it has been suggested that “Needing”ST, which depends on interoception and its prediction and its prediction error (i.e., difference between predicted need state and actual need state) within the anterior insula and mid‐posterior insula, respectively (Barrett & Simmons, 2015), also often recruits the anterior cingulate cortex (ACC) (Craig, 2003). Moreover, “Needing”ST related value attribution implicates the OFC (Balleine & O'Doherty, 2010; Ostlund & Balleine, 2007), while “Needing”ST related learning recruits the basolateral amygdala (Balleine & Killcross, 2006) and the long‐term association between external stimuli and their consequence on physiological need states recruits the caudate body and tail as well as the putamen, mostly the caudal‐ventral putamen (Amita et al., 2019; Kunimatsu et al., 2019; Schwabe & Wolf, 2010; Seger & Cincotta, 2005), and insular cortex (Balleine & Dickinson, 2000). Overall, previous functional magnetic resonance imaging (fMRI) meta‐analyses and studies on hunger in humans have revealed regions associated with sensory integration, reward processing, and taste, including the insula (Goldstone et al., 2009; Siep et al., 2009; van der Laan et al., 2011), the OFC (Führer et al., 2008; Goldstone et al., 2009; Siep et al., 2009; van der Laan et al., 2011), the amygdala (Führer et al., 2008; Goldstone et al., 2009; LaBar et al., 2001; Mohanty et al., 2008; van der Laan et al., 2011), the dorsal striatum (Siep et al., 2009; van der Laan et al., 2011), and the ACC (Führer et al., 2008; Goldstone et al., 2009; Siep et al., 2009), and many studies have found activations within the amygdala/parahippocampal gyrus (Chen & Zeffiro, 2020; LaBar et al., 2001; Mohanty et al., 2008). Hence, hunger can cautiously be used as a proxy for “Needing”ST. Thus, based on the inherent association between hunger (and thirst) and “Needing”ST, the insula and ACC, and to some extent the OFC, amygdala, and caudate may be engaged in the processing of “Needing”ST related value and contribute to directional value.
1.3. Two types of predictions and values
The conceptualization of “Wanting”ST and “Needing”ST here as processing of either wanted or needed stimuli implies two forms of predictions. Indeed, in both cases, the value of stimuli often depends on a prediction and a prediction error: in case of “Wanting”ST, that prediction is related to reward (unexpected reward or reward predicting cue) and is computed in the ventral striatum (Takahashi et al., 2016), while the prediction error is computed in the VTA (Schultz, 2015). For “Needing”ST, that prediction is related to interoception (predicted state vs. sensed state) and is said to be computed in visceromotor cortices (OFC, ACC, anterior insula), whereas the interoceptive prediction error is proposed to be computed within the mid‐posterior insula (Barrett & Simmons, 2015). Moreover, both “Wanting”ST and “Needing”ST establish a relation between the state (“wanting” state and “needing” state) and some external stimuli, where the state attributes some form of value to the stimuli. The value assigned to stimuli (by both “Wanting”ST and “Needing”ST) can have a directional effect or activational effect. The directional effect is linked to choice (preference or action selection) and directs towards or away from stimuli, while the activational effect is related to action and its initiation, maintenance, and vigor or effort (see Salamone et al., 2018). Indeed, “Wanting”ST depends on mesolimbic dopamine (Berridge, 1996) which provides full motivational value to reward as it provides both activational value (or effect) and directional value (or effect) to stimuli (see Salamone et al., 2018). “Needing”ST (by itself) does not seem to provide the activational value that “Wanting”ST provides to stimuli (Berridge, 2004; see also Salamone et al., 2018). However, “Needing”ST does provide directional value (Balleine, 1992; Salamone et al., 2018). Importantly, if both “Wanting”ST and “Needing”ST provide directional‐related value to stimuli that impacts choice, they do so in different ways. In the case of “Wanting”ST, that choice value is pavlovian cue triggered and stimulus related (Balleine, 2009; Berridge, 2012). In the case of “Needing”ST, that choice value is act‐outcome based (Balleine, 2009). Hence, depending on either state, stimulus value could be represented by different activation patterns in the brain (Berridge & Aldridge, 2009; Dayan & Balleine, 2002). However, no work has quantitatively tested this hypothesis.
Although “Wanting”ST takes both cues and physiological states, and the latter can be related to need states (Berridge, 1996; Zhang et al., 2009), not all physiological states are related to needs, some are related to emotions, drugs, and so forth (Berridge, 1996; Zhang et al., 2009). Moreover, need states can give and control value to relevant rewards in relation to their outcome by associating the discriminative properties of needs with the increased value placed on the reward (Balleine, 1992; Nader et al., 1997), and such “directing” motivational control through determination of specific outcome values is said to be dopamine independent (Balleine, 2005; Niv et al., 2006; see also Salamone et al., 2018). We believe there is an objective reaction to stimuli that are needed, even in absence of an incentive salience motivation, that is, “Wanting”ST. In the same way, there is an objective reaction to “Liking”ST in absence of “Wanting”ST (see Berridge, 1996); and in the same way, there is an objective “Wanting”ST, that can happen regardless of “Needing”ST or “Liking”ST (see Berridge, 2004). Regarding the difference between “Wanting”ST versus “Needing”ST, they likely provide two different “roles” (or values) to reward cues. We believe that, depending on situations or paradigms, “Wanting”ST related cues are stimuli that motivate action and bias behavior (see Robinson et al., 2014); whereas the “Needing”ST related cues are stimuli that are outcome relevant for the current need state (see Balleine, 2005), and our view is that those two roles/values do not necessarily apply at the same time to stimuli depending on situations/paradigms. Our study is thus about when needing a stimulus influences its processing without the wanting component for that stimulus, and when “Wanting”ST happens without “Needing”ST.
As discussed, neuroimaging studies in the activation pattern of “Wanting”ST related value shows consistent activation of the striatum, amygdala, and insula (Carter et al., 2009; Knutson et al., 2001, 2003; O'Doherty, 2004; O'Doherty et al., 2002; Oldham et al., 2018; Schott et al., 2008; Wilson et al., 2018). Those same regions have also been found in the activation pattern of “Needing”ST (Führer et al., 2008; Goldstone et al., 2009; LaBar et al., 2001; Mohanty et al., 2008; Siep et al., 2009; van der Laan et al., 2011). It is not clear how these regions contribute to either “Wanting”ST or “Needing”ST. Our goal is thus to use a meta‐analytic approach to compare the consistent brain activation patterns for “Wanting”ST and “Needing”ST related values by identifying similarities and differences between the brain activation patterns of these two states that guide value attribution and our consumption behaviors. To do this, we will quantitatively identify the consistent activation patterns after the observation of a reward cue/reward prediction (“Wanting”ST), versus while (or after) observing a food cue when hungry (“Needing”ST). We will then directly compare these activation patterns by conducting meta‐analytic conjunction and contrast analyses.
2. METHODOLOGY
We decided to use a meta‐analytic approach as it provides an opportunity to quantitatively assess brain activation patterns of “Wanting”ST versus “Needing”ST related values using large collections of data. This is useful as a summary of the existing literature is needed, not just because both concepts have rarely been directly compared and are often studied separately in neuroimaging studies, but also because each study might have low replicability, analytical and experimental flexibility, and/or small samples. Thus, our approach aims at identifying and comparing regions that are consistently activated for “Wanting”ST and those that are consistently activated for “Needing”ST. Specifically, we first conducted two meta‐analyses to quantitatively summarize results from fMRI published studies on the reward prediction for “Wanting”ST (activation maps taken when participants received a reward predicting cue that triggers reward seeking); and on perceiving food stimulus while being hungry for “Needing”ST (activation maps taken when participants perceived food while hungry). Second, we did a conjunction analysis to identify common regions that are consistently activated in both states. Finally, we contrasted “Wanting”ST and “Needing”ST consistent activation patterns by testing, [“Wanting”ST–“Needing”ST], and [“Needing”ST–“Wanting”ST].
2.1. Included articles
Based on the view that “Wanting”ST rests upon reward prediction that has been turned into a decision (see Berridge & Aldridge, 2009), we used the following keywords to identify articles related to “Wanting”ST:
((“prediction” AND “anticipation”) OR “desire” OR “wanting”)
Based on the view that “Needing”ST such as hunger depends on interoception (Craig, 2003) coming from deprivation of something biologically important, we used the following keywords to identify articles related to “Needing”ST:
(“alliesthesia” OR “interoceptive” OR “loss aversion” OR “need” OR “homeostasis” OR “modulating factor” OR “self‐specificity” OR “self‐referential” OR “hunger” OR “food deprivation”).
While the previous lists of keywords were specific to either “Wanting”ST or “Needing”ST, the following keywords were the same for both “Wanting”ST and “Needing”ST; those keywords were the following:
(“reward” OR “motivation” OR “goal directed” OR “decision‐making” OR “seeking” OR “incentive”) AND (“fMRI”)
These include words that are often conceptualized as related to “Wanting”ST and to “Needing”ST (Bouton, 2016; Panksepp, 2004).
For both “Wanting”ST or “Needing”ST, the following inclusion criteria were used: healthy subjects; whole‐brain analyses (with or without SVC), MNI or Talairach Coordinates (all Talairach coordinates were converted to MNI SPM152 in Ginger activation likelihood estimation [ALE] using Lancaster transform); maps were corrected (or cluster level corrected); activation contrast only.
With regard to “Wanting”ST, we typed the keywords on PubMed (February 2021). The database returned 159 articles. The main selection criteria were the presence of a cue that predicts a reward and triggers reward seeking contrasted with no prediction of reward (reward prediction > no reward prediction). After evaluation based on these criteria, 19 final articles were selected out of 26 that were fully read, and from which we found three additional articles from reviews and other articles that met all the criteria for “Wanting”ST for a total of 22 selected articles (see Table 2 for list of retained articles). Note that these rewards were mostly money or points, so they are not (directly) related to food, but they are used because “Wanting”ST or incentive motivation activates a general system, regardless of the type of stimulus (Bindra, 1968; Bouton, 2016). See PRISMA in Supporting Information for step‐by‐step exclusion of articles.
TABLE 2.
List of articles selected for ‘Wanting’ST selected articles
| Paper | Stimulus and cue | Task description | Contrast | Healthy participants |
|---|---|---|---|---|
| Schneider, M., Leuchs, L., Czisch, M., Sämann, P. G., & Spoormaker, V. I. (2018). Disentangling reward anticipation with simultaneous pupillometry/fMRI. NeuroImage, 178, 11–22. | Money, reward | Monetary Incentive Delay Task (MIDT) and Pupillometry | Reward anticipation‐control | 46 |
| Wu, C. C., Samanez‐Larkin, G. R., Katovich, K., & Knutson, B. (2014). Affective traits link to reliable neural markers of incentive anticipation. Neuroimage, 84, 279‐289. | Money | MIDT | Anticipation: Gain‐non Gain | 52 |
| Ubl, B., Kuehner, C., Kirsch, P., Ruttorf, M., Diener, C., & Flor, H. (2015). Altered neural reward and loss processing and prediction error signalling in depression. Social Cognitive and Affective Neuroscience, 10(8), 1102‐1112. | Money | Monetary reward paradigm | Anticipation: high gain vs control | 28 |
| Jia, T., Macare, C., Desrivières, S., Gonzalez, D. A., Tao, C., Ji, X., … Bokde, A. L. (2016). Neural basis of reward anticipation and its genetic determinants. Proceedings of the National Academy of Sciences, 113(14), 3879‐3884. | Money | MIDT | Anticipation: high win vs. no win | 1544 |
| Young, C. B., & Nusslock, R. (2016). Positive mood enhances reward‐related neural activity. Social Cognitive and Affective Neuroscience, 11(6), 934‐944. | Money | MIDT | Reward vs. nonreward (anticipation) | 40 |
| Millman, Z. B., Gallagher, K., Demro, C., Schiffman, J., Reeves, G. M., Gold, J. M., … & Buchanan, R. W. (2019). Evidence of reward system dysfunction in youth at clinical high‐risk for psychosis from two event‐related fMRI paradigms. Schizophrenia Research, 226, 111‐119. | Money | MIDT | Large received gain > small received gain | 41 |
| Navas, J. F., Barrós‐Loscertales, A., Costumero‐Ramos, V., Verdejo‐Román, J., Vilar‐López, R., & Verdejo‐García, A. (2018). Excessive body fat linked to blunted somatosensory cortex response to general reward in adolescents. International Journal of Obesity, 42(1), 88. | Money | MIDT | Reward anticipation | 68 |
| Herbort, M. C., Soch, J., Wüstenberg, T., Krauel, K., Pujara, M., Koenigs, M., … & Schott, B. H. (2016). A negative relationship between ventral striatal loss anticipation response and impulsivity in borderline personality disorder. NeuroImage: Clinical, 12, 724‐736. | Money | MIDT | Gain anticipation | 23 |
| Kohls, G., Perino, M. T., Taylor, J. M., Madva, E. N., Cayless, S. J., Troiani, V., … Schultz, R. T. (2013). The nucleus accumbens is involved in both the pursuit of social reward and the avoidance of social punishment. Neuropsychologia, 51(11), 2062‐2069. | Social incentive | SIDT | Anticipation of social approval | 22 |
| Kumar, P., Berghorst, L. H., Nickerson, L. D., Dutra, S. J., Goer, F. K., Greve, D. N., & Pizzagalli, D. A. (2014). Differential effects of acute stress on anticipatory and consummatory phases of reward processing. Neuroscience, 266, 1‐12. | Money | MIDT | Anticipation (Reward vs. No‐incentive Cue) | 18 |
| Bradley, K. A., Case, J. A., Freed, R. D., Stern, E. R., & Gabbay, V. (2017). Neural correlates of RDoC reward constructs in adolescents with diverse psychiatric symptoms: A Reward Flanker Task pilot study. Journal of Affective Disorders, 216, 36‐45. | Money | Reward flanker task | Reward anticipation vs. implicit baseline | 22 |
| Richter, A., Petrovic, A., Diekhof, E. K., Trost, S., Wolter, S., & Gruber, O. (2015). Hyperresponsivity and impaired prefrontal control of the mesolimbic reward system in schizophrenia. Journal of Psychiatric Research, 71, 8‐15. | Points, targets and CS | Desire‐reason paradigm | Desire context | 16 |
| Reason context | ||||
| Gluth, S., Rieskamp, J., & Büchel, C. (2013). Neural evidence for adaptive strategy selection in value‐based decision‐making. Cerebral Cortex, 24(8), 2009‐2021. | Investment | Dynamic learning task | Expected value | 24 |
| Trost, S., Diekhof, E. K., Mohr, H., Vieker, H., Krämer, B., Wolf, C., … Gruber, O. (2016). Investigating the impact of a genome‐wide supported bipolar risk variant of MAD1L1 on the human reward system. Neuropsychopharmacology, 41(11), 2679. | Points, targets and CS | Desire‐reason paradigm | Desire context | 224 |
| Reason context | ||||
| Trost, S., Diekhof, E. K., Zvonik, K., Lewandowski, M., Usher, J., Keil, M., … Gruber, O. (2014). Disturbed anterior prefrontal control of the mesolimbic reward system and increased impulsivity in bipolar disorder. Neuropsychopharmacology, 39(8), 1914. | Points, targets and CS | Desire‐reason paradigm | Desire context | 16 |
| Yu, R., Mobbs, D., Seymour, B., Rowe, J. B., & Calder, A. J. (2014). The neural signature of escalating frustration in humans. Cortex, 54, 165‐178. ISO 690 | Cue and coin | Multitrial reward schedule task | Cue–block | 27 |
| Cue (increased proximity) | ||||
| Cue (increased expended effort) | ||||
| Krebs, R. M., Schott, B. H., Schütze, H., & Düzel, E. (2009). The novelty exploration bonus and its attentional modulation. Neuropsychologia, 47(11), 2272‐2281. | cue, reward | Number comparison task (NCT) | Reward‐predicting cues in Exp 1: Contrast reward vs. neutral | 24 Exp1 |
| 20 Exp 2 | ||||
| Familiar reward‐predicting cues Exp 1 | ||||
| Reward‐predicting cues in Exp 2: Contrast reward vs. neutral | ||||
| Novel reward‐predicting cues Exp 2 | ||||
| Articles from other sources | ||||
| Samanez‐Larkin, G. R., Gibbs, S. E., Khanna, K., Nielsen, L., Carstensen, L. L., & Knutson, B. (2007). Anticipation of monetary gain but not loss in healthy older adults. Nature Neuroscience, 10(6), 787. | Money | MIDT | Gain vs. nongain anticipation: younger | 12 |
| 12 | ||||
| Gain vs. nongain anticipation: older | ||||
| Simon, J. J., Walther, S., Fiebach, C. J., Friederich, H. C., Stippich, C., Weisbrod, M., & Kaiser, S. (2010). Neural reward processing is modulated by approach‐and avoidance‐related personality traits. Neuroimage, 49(2), 1868‐1874. | Money | MIDT | Anticipation of reward vs. nonreward | 24 |
| Wittmann, B. C., Schott, B. H., Guderian, S., Frey, J. U., Heinze, H. J., & Düzel, E. (2005). Reward‐related FMRI activation of dopaminergic midbrain is associated with enhanced hippocampus‐dependent long‐term memory formation. Neuron, 45(3), 459‐467. | Money | MIDT | Reward anticipation | 16 |
| Murray, L., Lopez‐Duran, N. L., Mitchell, C., Monk, C. S., & Hyde, L. W. (2020). Neural mechanisms of reward and loss processing in a low‐income sample of at‐risk adolescents. Social Cognitive and Affective Neuroscience, 15(12), 1299‐1314. | Points | Lottery choice task | Reward anticipation > neutral anticipation | 128 |
| Yao, Y. W., Liu, L., Worhunsky, P. D., Lichenstein, S., Ma, S. S., Zhu, L., … Yip, S. W. (2020). Is monetary reward processing altered in drug‐naïve youth with a behavioral addiction? Findings from internet gaming disorder. NeuroImage: Clinical, 26, 102202. | Money | MID task | Gain anticipation | 27 |
Regarding “Needing”ST related articles, we typed the keywords on PubMed (February 2021). The database returned 376 articles. The main logic was to select experiments when subjects were in a hungry state and perceiving a food stimulus. We looked for both “hunger > baseline” as well as “hunger > satiety” contrasts, because of the inherent subtraction logic of fMRI and in order to have a larger number of experiments. Hence, the two main criteria were: (1) presence of a privation contrast: hunger + stimulus > satiety + stimulus, or hunger + stimulus > baseline; (2) the participant was perceiving some food stimulus which could be presented in any modality: visual, taste, odor, and so forth. Using the selection criteria (see Table 1 for all criteria), we kept 26 articles. After fully reading the final 26 articles, nine were selected, and we found some additional ones through other articles and reviews, and seven among them matched all criteria for “Needing”ST (hunger) for a total of 16 articles (see Table 3 for list of retained articles). See PRISMA and Supporting Information for step‐by‐step exclusion of articles.
TABLE 1.
Selection criteria
| Criteria | Needs (hunger) | “Wanting” |
|---|---|---|
| Privation contrast | Yes | N/A |
| Presence of cue indicating the reward | Yes | Yes |
| Cue that triggers decision (to get the reward to be gained during the task) | Not necessarily | Yes |
| fMRI contrast taken only during the anticipation (cue) or after (during) the viewing of the cue | Yes | Yes |
| The reward is relevant for the need (self‐specific) | Yes | N/A |
| Do not contrast two rewards | N/A (e.g. high calorie–low calorie experiments were included) | Yes (with one exception) |
| Healthy individuals only | Yes (however, we took people that had up to 30–35 of BMI, especially in contrasts when healthy and overweight were mixed) | Yes |
| MNI or Talairach Coordinates | Yes | Yes |
| Whole brain contrast (with or without SVC) | Yes | Yes |
| Corrected | Yes | Yes |
| Activation contrast only | Yes | Yes |
| Excluded: MRI and resting states; cognitive conjunction analysis; and functional connectivity results | Yes | Yes |
Note: The red colored “yes” means the criterion is crucial for the definition of either “Wanting”ST or “Needing”ST. Most mean BMI values were below 30, which is the threshold of obesity as defined by the World Health Organization (World Health Organization, 2020); however, we took people that had up to 30–35 of BMI, especially in contrast when healthy and overweight were mixed. For the Millman et al.’s (2020) study, we took the contrast of “large gain > small gain” (so exceptionally, we contrasted two rewards); first because there were no contrast for gain alone in general, and because small gains as well as large losses were received when participants failed to respond within the allowable time window, so these two outcomes served as de facto negative RPEs.
Abbreviation: MRI, magnetic resonance imaging.
TABLE 3.
List of articles selected for ‘Needing’ST
| Paper | Contrast | Stimuli | Task | Healthy Participants | Fasted hours |
|---|---|---|---|---|---|
| Jiang, T., Soussignan, R., Schaal, B., & Royet, J. P. (2014). Reward for food odors: An fMRI study of liking and wanting as a function of metabolic state and BMI. Social Cognitive and Affective Neuroscience, 10(4), 561‐568. | Liking–Wanting (hunger) | Odor | Odor presentation and rating | 12 | |
| Wanting – Liking (hunger) | |||||
| Food–NFood (hunger) | |||||
| Green, E., Jacobson, A., Haase, L., & Murphy, C. (2015). Neural correlates of taste and pleasantness evaluation in the metabolic syndrome. Brain Research, 1620, 57‐71. | Hunger–satiety: Control | Taste | Swallowing aqueous solution | 15 | 12 h |
| Sucrose > Caffeine (during hunger) | |||||
| Caffeine > Sucrose (hunger) | |||||
| Martens, M. J., Born, J. M., Lemmens, S. G., Karhunen, L., Heinecke, A., Goebel, R., … Westerterp‐Plantenga, M. S. (2013). Increased sensitivity to food cues in the fasted state and decreased inhibitory control in the satiated state in the overweight. The American Journal of Clinical Nutrition, 97(3), 471‐479. | Fasted: F > NF | Visual | Viewing food and nonfood pictures | 40 | 10 h |
| Fasted: stimuli–subject group | |||||
| Fasted: correlation F > NF with BMI | |||||
| LaBar, K. S., Gitelman, D. R., Parrish, T. B., Kim, Y. H., Nobre, A. C., & Mesulam, M. (2001). Hunger selectively modulates corticolimbic activation to food stimuli in humans. Behavioral Neuroscience, 115(2), 493. | Hungry–satiated | Visual | Viewing food and tool images | 17 | 8 h |
| Harding, I. H., Andrews, Z. B., Mata, F., Orlandea, S., Martinez‐Zalacain, I., Soriano‐Mas, C., … Verdejo‐Garcia, A. (2018). Brain substrates of unhealthy versus healthy food choices: influence of homeostatic status and body mass index. International Journal of Obesity, 42(3), 448. | Healthy vs. unhealthy food choice: fasted > satiated | Visual | Participants were asked to select an option using a two‐button response box | 30 | 10 h |
| Frank, S., Laharnar, N., Kullmann, S., Veit, R., Canova, C., Hegner, Y. L., … Preissl, H. (2010). Processing of food pictures: Influence of hunger, gender and calorie content. Brain Research, 1350, 159‐166. | HiCal hungry vs. HiCal satiated | Visual | One‐back task: press a button, either to indicate that the seen image was the same or another button to indicate that the picture was not the same | 12 | 8 h |
| Holsen, L. M., Zarcone, J. R., Thompson, T. I., Brooks, W. M., Anderson, M. F., Ahluwalia, J. S., & Savage, C. R. (2005). Neural mechanisms underlying food motivation in children and adolescents. Neuroimage, 27(3), 669‐676. | Premeal: Food > nonfood | Visual | Viewing pictures of food, animals, and baseline control images | 9 | 4 h |
| Jacobson, A., Green, E., Haase, L., Szajer, J., & Murphy, C. (2019). Differential effects of bmi on brain response to odor in olfactory, reward and memory regions: Evidence from fMRI. Nutrients, 11(4), 926. | Odor during the hunger condition | Odor/taste | Odor stimuli delivered to the tongue | 40 | 12 h |
| Articles from other sources | |||||
| Haase, L., Green, E., & Murphy, C. (2011). Males and females show differential brain activation to taste when hungry and sated in gustatory and reward areas. Appetite, 57(2), 421‐434. | Hunger × male × Sucrose > water | Taste | Taste stimuli presentation | 21 | 12 h |
| Hunger × female × NaCl > water | |||||
| Hunger × female × Caffeine > water | |||||
| Hunger × female × Sucrose > water | |||||
| Hunger × female × citric acid > water | |||||
| Führer, D., Zysset, S., & Stumvoll, M. (2008). Brain activity in hunger and satiety: An exploratory visually stimulated FMRI study. Obesity, 16(5), 945‐950. | Hunger > Satiety | Visual | Viewing pictures and two‐back task: the subject was asked to press a button when the same letter was shown two steps earlier | 12 | 14 h |
| Haase, L., Cerf‐Ducastel, B., & Murphy, C. (2009). Cortical activation in response to pure taste stimuli during the physiological states of hunger and satiety. Neuroimage, 44(3), 1008‐1021. | Hunger > satiety × sucrose | Taste | Stimulus presentation delivered to the tip of the tongue | 18 | 12 h |
| Hunger > satiety × caffeine | |||||
| Hunger > satiety × saccharin | |||||
| Hunger > satiety × citric acid | |||||
| Uher, R., Treasure, J., Heining, M., Brammer, M. J., & Campbell, I. C. (2006). Cerebral processing of food‐related stimuli: Effects of fasting and gender. Behavioural Brain Research, 169(1), 111‐119. | Fasting > satiety × food‐related stimuli (chocolate and chicken) | Visual | Viewing photographs of food | 18 | 24 h |
| Fasting > satiety × food‐related stimuli (chocolate) | |||||
| Fasting > satiety × food‐related stimuli (Chicken) | |||||
| Holsen, L. M., Zarcone, J. R., Brooks, W. M., Butler, M. G., Thompson, T. I., Ahluwalia, J. S., … Savage, C. R. (2006). Neural mechanisms underlying hyperphagia in Prader‐Willi syndrome. Obesity, 14(6), 1028‐1037. | HW × premeal × food > non‐food | Visual | Viewing pictures of food, animals, and control | 9 | 4 h |
| Hw × premeal × nonfood > food | |||||
| Jacobson, A., Green, E., & Murphy, C. (2010). Age‐related functional changes in gustatory and reward processing regions: An fMRI study. Neuroimage, 53(2), 602‐610. | Hunger × older adults × sucrose | Taste | Stimuli were delivered orally | 38 | 12 h |
| Hunger × young adults × sucrose | |||||
| Hunger × older adults × citric acid | |||||
| Hunger × younger adults × citric acid | |||||
| Hunger × older adults × NaCl | |||||
| Hunger × younger adults × NaCl | |||||
| Hunger × older adults × caffeine | |||||
| Hunger × younger adults × caffeine | |||||
| Cheah, Y. S., Lee, S., Ashoor, G., Nathan, Y., Reed, L. J., Zelaya, F. O., … Amiel, S. A. (2014). Ageing diminishes the modulation of human brain responses to visual food cues by meal ingestion. International Journal of Obesity, 38(9), 1186. | Fasted > fed × visual food cue | Visual | Viewing food and nonfood | 24 | 8 h |
| He, Q., Huang, X., Zhang, S., Turel, O., Ma, L., & Bechara, A. (2019). Dynamic causal modeling of insular, striatal, and prefrontal cortex activities during a food‐specific Go/NoGo task. Biological Psychiatry: Cognitive Neuroscience and Neuroimaging, 4(12), 1080‐1089. | Hungry > satiated | Visual | Food pictures | 45 | 14–15 h |
In order to more easily disentangle “Wanting”ST versus “Needing”ST, the experiments included in “Wanting”ST did not include need states, and the “Needing”ST related experiments did not include situations in which a cue triggers behavior/reward seeking. This might be viewed as nonfasting versus fasting, but such interpretation should be taken cautiously as it is not about the same reward type, and “Wanting”ST and “Needing”ST are dependent on different situations/paradigms and reward cue roles. Of note, “Wanting”ST studies did not explicitly exclude food‐related studies and were not limited to secondary rewards. However, our selection criteria resulted in the fact that we did not find food related “Wanting”ST studies to include. Hence, unintentionally, “Needing”ST included primary reward whereas “Wanting”ST included secondary rewards and motor action. For “Needing”ST, the included studies used a contrast on perception of food‐related stimuli while hungry versus while satiated. Thus, brain activity elicited by food itself would cancel out, and only the “Needing”ST part should remain. Regarding the experimental task, we share the view that “Wanting”ST is related to the preparatory and motivational excitement for motor behavior, specifically within the NAcc (Cardinal et al., 2002). Aside from that, the “Wanting”ST contrast is between anticipation of a reward cue versus anticipation of nonreward cues, where participants responded whether they were rewarded or not. In that sense, motor preparation per se would likely cancel out or would not account for all the brain regions of such contrast.
2.2. Meta‐analyses
Meta‐analyses were conducted with the activation likelihood estimation (ALE) approach using the Brainmap's GingerALE application. Independently introduced by Turkeltaub et al. (2002) and by Chein et al. (2002) and revised by Eickhoff et al. (2009), the ALE meta‐analysis treats activation foci not as single point, but as spatial probability distributions that are centered at the given coordinates (Eickhoff et al., 2012). The Eickhoff et al.’s (2009) revised ALE algorithm models the spatial uncertainty by using an estimation of the intersubject and interlaboratory variability (which is typically observed in neuroimaging experiments). Then, union of activation probabilities for each voxel of all included experiment is computed to give an ALE map, and a permutation procedure (in which datasets are created similar to the real data in terms of number of experiments, foci per experiments and number of subjects, but in which foci are randomly distributed) is used in order to test the differentiation between true convergence of foci and random clustering (Eickhoff et al., 2012). As a method of inference, the new algorithm uses random‐effects analysis that calculates the above‐chance clustering between experiments. Furthermore, the new algorithm gives more weight to gray matter compared to white matter by limiting the meta‐analysis to an anatomically constrained space specified by a gray matter mask. Contrast analyses are based on two different datasets (i.e., two previous ALE results) and thus compare two different sets of foci for statistically significant differences, and the conjunction is the intersection of the thresholded maps.
In our analyses, we used the MNI152 coordinate system and the less conservative (larger) mask size. For “Wanting”ST, there were 21 articles, 34 experiments, 3306 subjects, and 572 foci (see Tables 2 and 3 for all included articles). For “Needing”ST, (hunger), we had 16 articles, 38 experiments, 733 subjects, and 494 foci. In our study, for main individual meta‐analyses, all maps were thresholded using a cluster‐level family‐wise error (cFWE) correction (p < .05) with a cluster‐forming threshold of p < .001(uncorrected at the voxel level) (Eklund et al., 2016; Woo et al., 2014) and 1000 permutations. For the contrast meta‐analyses, we used the two cFWE corrected maps with p < .01 (uncorrected at the voxel level), 10,000 permutations (see Eickhoff et al., 2011), and the conjunction was the intersection of the two cFWE thresholded maps. Maps from meta‐analyses were overlaid on a MNI template and viewed using Mango (http://ric.uthscsa.edu/mango/).
Our inclusion criteria (such as including only corrected results and experiments) lowered the number of included experiments. Thus, to confirm that our main meta‐analytic results were not driven by the coordinates from a single publication, we conducted validation analyses using a leave‐one‐experiment‐out (LOEO) approach. In this approach, on each fold, one contrast (i.e., experiment) was excluded, and the ALE meta‐analysis was conducted on the remaining N–1 contrasts. Thus, the results from this procedure consisted of brain regions that were identified in every fold of the LOEO and are not mainly driven by a single contrast.
3. RESULTS
3.1. Main meta‐analyses
3.1.1. “Wanting”ST
Our first meta‐analysis was on “Wanting”ST (Table 4 and Figure 1). This meta‐analysis revealed consistent activations within the following regions: the left putamen, the left globus pallidus (which encompassed the nucleus accumbens), the left caudate body and right caudate head, the left substantia nigra, the right red nucleus (encompassing the ventral tegmental area), the right hypothalamus, the bilateral thalamus, the left precentral gyrus, the left inferior parietal lobule, the right dorso‐lateral and medial prefrontal cortex, the right superior parietal lobule, the right claustrum (whose cluster was mainly the anterior insula). Of note, because we had much more MID (monetary incentive delay) tasks (see Knutson et al., 2000) in the “Wanting”ST contrast, we conducted a single meta‐analysis with only studies that did not use the MID task (please see non‐MID task meta‐analysis in the Supporting Information), and we found peak activity within the ventral and dorsal striatum, the dopaminergic midbrain (VTA/SN), and anterior insula. Thus, though experimental tasks for “Wanting”ST included a lot of MID tasks, other included paradigms elicited (separately from the MID tasks) the same mesolimbic dopamine and ventral striatal network that has been related to incentive salience “Wanting”ST; although in terms of overall whole brain pattern, they might have differed.
TABLE 4.
Coordinates for peak activated clusters in the ‘Wanting’ST condition
| Cluster # | x | y | z | ALE | P | Z | Label (Nearest Gray Matter within 5 mm) |
|---|---|---|---|---|---|---|---|
| 1 | −16 | 8 | 2 | 0.073149 | 8.39E‐21 | 9.281936 | Left Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 1 | −32 | 18 | 4 | 0.072856 | 1.07E‐20 | 9.256376 | Left Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 1 | 12 | 8 | 0 | 0.062949 | 3.04E‐17 | 8.364339 | Right Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Head |
| 1 | −18 | 0 | 14 | 0.041655 | 1.70E‐10 | 6.279265 | Left Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Body |
| 1 | 18 | −6 | 8 | 0.023581 | 1.41E‐05 | 4.186929 | Right Cerebrum.Sub‐lobar.Thalamus.Gray Matter.Ventral Anterior Nucleus |
| 1 | −10 | −6 | −2 | 0.022746 | 2.26E‐05 | 4.078712 | Left Cerebrum.Sub‐lobar.Thalamus.Gray Matter.* |
| 1 | 10 | −6 | −6 | 0.022589 | 2.48E‐05 | 4.057735 | Right Cerebrum.Sub‐lobar.*.Gray Matter.Hypothalamus |
| 1 | −16 | −4 | −2 | 0.019475 | 1.37E‐04 | 3.639444 | Left Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Medial Globus Pallidus |
| 1 | 8 | −6 | 10 | 0.018669 | 2.11E‐04 | 3.525875 | Right Cerebrum.Sub‐lobar.Thalamus.Gray Matter.Anterior Nucleus |
| 2 | −6 | −22 | −18 | 0.070495 | 7.37E‐20 | 9.047366 | Left Brainstem.Midbrain.*.Gray Matter.Substania Nigra |
| 2 | 6 | −22 | −18 | 0.062512 | 4.28E‐17 | 8.323777 | Right Brainstem.Midbrain.*.Gray Matter.Red Nucleus |
| 3 | −6 | 8 | 52 | 0.055675 | 7.87E‐15 | 7.681671 | Left Cerebrum.Frontal Lobe.Medial Frontal Gyrus.Gray Matter.Brodmann area 6 |
| 4 | 34 | 22 | 2 | 0.071349 | 3.69E‐20 | 9.122826 | Right Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 5 | −28 | −8 | 56 | 0.036814 | 4.27E‐09 | 5.757467 | Left Cerebrum.Frontal Lobe.Precentral Gyrus.Gray Matter.Brodmann area 6 |
| 6 | −44 | −36 | 46 | 0.02985 | 3.47E‐07 | 4.963281 | Left Cerebrum.Parietal Lobe.Inferior Parietal Lobule.Gray Matter.Brodmann area 40 |
| 7 | 38 | 36 | 28 | 0.045997 | 8.58E‐12 | 6.728507 | Right Cerebrum.Frontal Lobe.Middle Frontal Gyrus.Gray Matter.Brodmann area 9 |
| 8 | 28 | −4 | 50 | 0.03858 | 1.34E‐09 | 5.950417 | Right Cerebrum.Frontal Lobe.Middle Frontal Gyrus.Gray Matter.Brodmann area 6 |
| 9 | 34 | −52 | 44 | 0.0408 | 3.04E‐10 | 6.188297 | Right Cerebrum.Parietal Lobe.Superior Parietal Lobule.Gray Matter.Brodmann area 7 |
| 10 | −44 | 2 | 34 | 0.045089 | 1.62E‐11 | 6.635556 | Left Cerebrum.Frontal Lobe.Precentral Gyrus.Gray Matter.Brodmann area 6 |
| 11 | −28 | 38 | 12 | 0.027964 | 1.09E‐06 | 4.736878 | No Gray Matter found |
| 11 | −32 | 44 | 14 | 0.024484 | 8.43E‐06 | 4.302969 | Left Cerebrum.Frontal Lobe.Middle Frontal Gyrus.Gray Matter.Brodmann area 10 |
| 11 | −36 | 50 | 22 | 0.019078 | 1.69E‐04 | 3.583664 | Left Cerebrum.Frontal Lobe.Middle Frontal Gyrus.Gray Matter.Brodmann area 9 |
| 12 | 46 | −34 | 46 | 0.035456 | 1.02E‐08 | 5.608079 | Right Cerebrum.Parietal Lobe.Inferior Parietal Lobule.Gray Matter.Brodmann area 40 |
Abbreviation: ALE, activation likelihood estimation.
FIGURE 1.
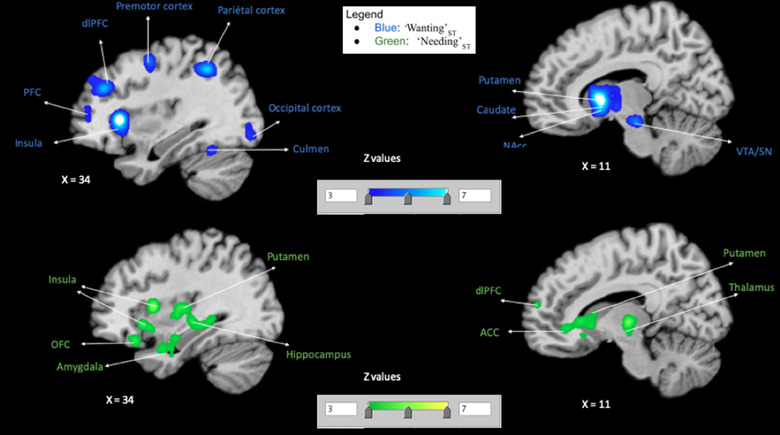
Single meta‐analyses maps. Maps for activated clusters in each condition: “Wanting”ST (blue) and “Needing”ST (green), showing activation pattern for each
3.1.2. “Needing”ST
Next, we conducted an individual meta‐analysis on “Needing”ST (hunger with stimulus; Table 5 and Figure 1). This second meta‐analysis revealed consistent activations in: the bilateral anterior insula, right middle and posterior insula, right thalamus, left claustrum, right hippocampus, bilateral putamen, right caudate body, right caudate head (encompassing the NAcc), and right posterior putamen (encompassing the caudate tail), amygdala, bilateral anterior cingulate area (encompassing the right OFC), right uncus and left subcallosal area (which can be considered as entorhinal cortex (Fischl et al., 2009), and the right mammillary body.
TABLE 5.
Coordinates for peak activated clusters in the ‘Needing’ST condition
| Cluster # | x | y | z | ALE | P | Z | Label (nearest gray matter within 5 mm) |
|---|---|---|---|---|---|---|---|
| 1 | 38 | 10 | 6 | 0.030085 | 9.63E‐09 | 5.61851 | Right Cerebrum.Sub‐lobar.Insula.Gray Matter.Brodmann area 13 |
| 1 | 14 | −16 | 0 | 0.027834 | 5.17E‐08 | 5.32083 | Right Cerebrum.Sub‐lobar.Thalamus.Gray Matter.Mammillary Body |
| 1 | 42 | −4 | −2 | 0.022411 | 2.42E‐06 | 4.571283 | Right Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 1 | 44 | 2 | −10 | 0.022346 | 2.53E‐06 | 4.56251 | Right Cerebrum.Sub‐lobar.Insula.Gray Matter.Brodmann area 13 |
| 1 | 48 | −12 | 4 | 0.021699 | 3.97E‐06 | 4.467 | Right Cerebrum.Sub‐lobar.Insula.Gray Matter.Brodmann area 13 |
| 1 | 32 | −38 | −4 | 0.021315 | 5.15E‐06 | 4.410615 | Right Cerebrum.Temporal Lobe.Sub‐Gyral.Gray Matter.Hippocampus |
| 1 | 34 | −12 | 6 | 0.020947 | 6.64E‐06 | 4.355373 | Right Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 1 | 34 | 18 | −8 | 0.020895 | 6.92E‐06 | 4.346396 | Right Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 1 | 36 | −22 | −4 | 0.020609 | 8.37E‐06 | 4.304496 | Right Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 1 | 28 | −8 | 0 | 0.019476 | 1.78E‐05 | 4.134444 | Right Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 1 | 30 | −50 | −12 | 0.016798 | 9.81E‐05 | 3.723894 | Right Cerebellum.Anterior Lobe.Culmen.Gray Matter.* |
| 2 | 14 | 36 | −4 | 0.022022 | 3.18E‐06 | 4.514158 | Right Cerebrum.Limbic Lobe.Anterior Cingulate.Gray Matter.Brodmann area 24 |
| 2 | 10 | 24 | 0 | 0.021439 | 4.75E‐06 | 4.428455 | Right Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Head |
| 2 | 10 | 14 | −2 | 0.017611 | 5.89E‐05 | 3.850592 | Right Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Head |
| 2 | 8 | 20 | −10 | 0.017451 | 6.52E‐05 | 3.825874 | Right Cerebrum.Limbic Lobe.Anterior Cingulate.Gray Matter.Brodmann area 25 |
| 2 | 12 | 16 | 4 | 0.017117 | 8.02E‐05 | 3.774332 | Right Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Body |
| 2 | 14 | 12 | 8 | 0.016643 | 1.08E‐04 | 3.698454 | Right Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Body |
| 2 | 14 | 22 | −12 | 0.015113 | 2.87E‐04 | 3.443822 | Right Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Head |
| 3 | −38 | 0 | −2 | 0.030925 | 5.07E‐09 | 5.728328 | Left Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 3 | −40 | 8 | 12 | 0.017386 | 6.77E‐05 | 3.816551 | Left Cerebrum.Sub‐lobar.Insula.Gray Matter.Brodmann area 13 |
| 4 | 26 | 6 | −16 | 0.021686 | 3.99E‐06 | 4.465524 | Right Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 4 | 30 | −10 | −18 | 0.02053 | 8.83E‐06 | 4.292494 | Right Cerebrum.Limbic Lobe.Parahippocampal Gyrus.Gray Matter.Amygdala |
| 4 | 32 | −4 | −26 | 0.019465 | 1.80E‐05 | 4.131432 | Right Cerebrum.Limbic Lobe.Parahippocampal Gyrus.Gray Matter.Amygdala |
| 4 | 34 | 4 | −28 | 0.019396 | 1.87E‐05 | 4.122383 | Right Cerebrum.Limbic Lobe.Uncus.Gray Matter.Brodmann area 28 |
| 5 | −14 | 18 | −20 | 0.020973 | 6.55E‐06 | 4.358362 | Left Cerebrum.Frontal Lobe.Medial Frontal Gyrus.Gray Matter.Brodmann area 25 |
| 5 | −26 | 4 | −16 | 0.018197 | 4.06E‐05 | 3.940716 | Left Cerebrum.Frontal Lobe.Subcallosal Gyrus.Gray Matter.Brodmann area 34 |
| 5 | −14 | 22 | −12 | 0.017613 | 5.89E‐05 | 3.850592 | Left Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Head |
| 5 | −18 | 10 | −18 | 0.016592 | 1.12E‐04 | 3.690466 | Left Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 5 | −6 | 22 | −10 | 0.015431 | 2.34E‐04 | 3.498593 | Left Cerebrum.Limbic Lobe.Anterior Cingulate.Gray Matter.Brodmann area 24 |
| 6 | −34 | −22 | −4 | 0.025242 | 3.35E‐07 | 4.970016 | Left Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
Abbreviation: ALE, activation likelihood estimation.
3.2. Validation results (LOEO analyses)
The key output from the LOEO analysis was related to the robustness per cluster. That is, in what probability percentage a given cluster was observed. Here, we show from the LOEO analysis brain regions that have 100% probability of being activated in all experiments included in the meta‐analyses.
3.2.1. “Wanting”ST (Supporting Information and Figure 2)
FIGURE 2.
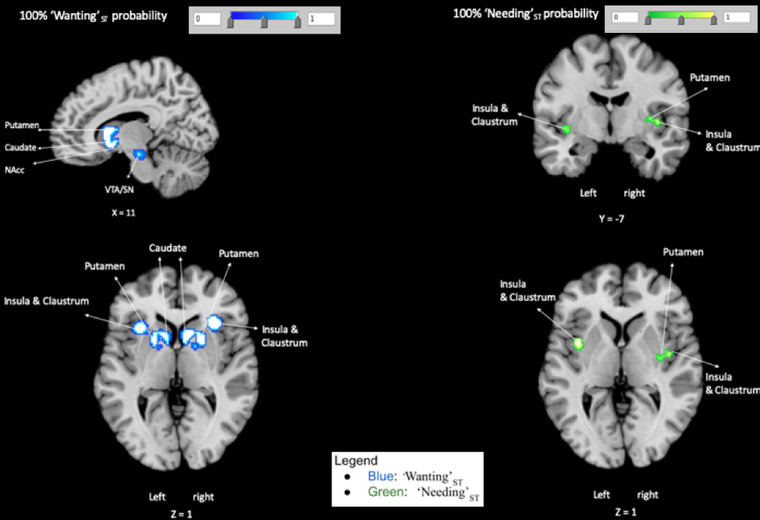
100% Probability maps. LOEO Maps for clusters of “Wanting”ST (blue) activation pattern and “Needing”ST (green) activation pattern that have 100% probability of being activated in each included experiments
For “Wanting”ST, consistent activations were identified in ALE‐LOEO with 100% probability in the following peak regions: right midbrain (VTA and SN), right putamen (that included the caudate and the NAcc), left ACC, left caudate, left OFC, left anterior insula, and left inferior parietal lobule.
3.2.2. “Needing”ST (Supporting Information and Figure 2)
Regarding, “Needing”ST ALE‐LOEO meta‐analysis revealed three peaks with 100% consistent activations in all experiments, within the bilateral middle insula, that included the caudoventral putamen and the claustrum.
3.3. Conjunction and contrasts meta‐analyses
Contrasts and conjunction analyses were based on ALE results of the two previous ALE results (“Wanting”ST AND “Needing”ST) that were compared for statistically significant differences and similarities.
3.3.1. “Wanting”ST AND “Needing”ST conjunction
The conjunction between “Wanting”ST AND “Needing”ST resulted in consistent activations within the head and body of the right caudate nucleus (the activated region does not include the nucleus accumbens), right claustrum, and right anterior insula (Table 6 and Figure 3).
TABLE 6.
Coordinates for peak activated clusters in the ‘Wanting’ST AND ‘Needing’ST conjunction
| Cluster # | x | y | z | ALE | Label (nearest gray matter within 5 mm) |
|---|---|---|---|---|---|
| 1 | 10 | 14 | −2 | 0.017611 | Right Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Head |
| 1 | 12 | 16 | 4 | 0.017117 | Right Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Body |
| 1 | 14 | 12 | 8 | 0.016643 | Right Cerebrum.Sub‐lobar.Caudate.Gray Matter.Caudate Body |
| 2 | 34 | 20 | −6 | 0.020654 | Right Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 3 | 36 | 16 | 6 | 0.013647 | Right Cerebrum.Sub‐lobar.Insula.Gray Matter.Brodmann area 13 |
Abbreviation: ALE, activation likelihood estimation.
FIGURE 3.
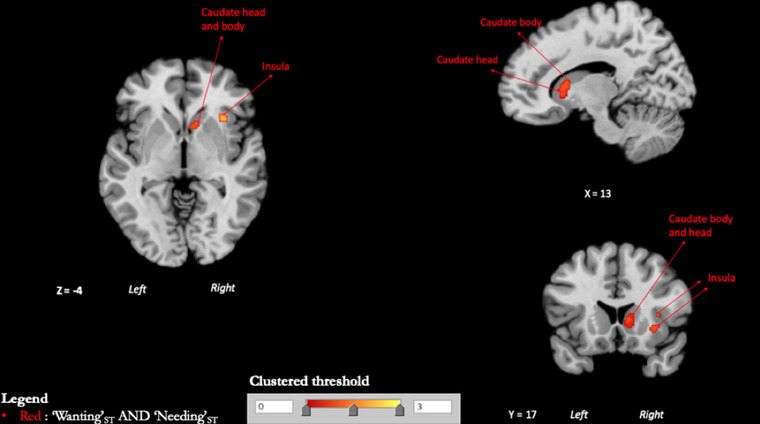
Conjunction maps. Clustered thresholded maps showing the intersection between activation patterns of “Wanting”ST and “Needing”ST
3.3.2. Contrast: “Wanting”ST–“Needing”ST
Compared to “Needing”ST, “Wanting”ST more consistently activated regions of the left lateral globus pallidus (which encompassed the nucleus accumbens), the left red nucleus (encompassing the ventral tegmental area), right substantia nigra (SN), bilateral putamen, left anterior insula, the left precentral gyrus, the right superior parietal lobule, the left inferior parietal lobule, the right claustrum, the left anterior dorsolateral prefrontal cortex, and the right angular gyrus (Table 7 and Figure 4).
TABLE 7.
Coordinates for peak activated clusters in the ‘Wanting’ST–‘Needing’ST contrast
| Cluster # | x | y | z | P | Z | Label (Nearest gray matter within 5 mm) |
|---|---|---|---|---|---|---|
| 1 | −16.4 | 9.4 | 3.2 | 0 | 3.890594 | Left Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 1 | −19.3 | 6 | 5.3 | 1.00E‐04 | 3.719017 | Left Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 1 | −27.3 | 22 | 6.7 | 2.00E‐04 | 3.540084 | Left Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 1 | −32.4 | 19.2 | 4.8 | 3.00E‐04 | 3.431614 | Left Cerebrum.Sub‐lobar.Insula.Gray Matter.Brodmann area 13 |
| 2 | 15 | 5.5 | −2.5 | 0 | 3.890594 | Right Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Lateral Globus Pallidus |
| 2 | 21 | 1 | 9 | 3.00E‐04 | 3.431614 | Right Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 3 | −0.7 | −23 | −19.1 | 0 | 3.890594 | Left Brainstem.Midbrain.*.Gray Matter.Red Nucleus |
| 3 | 8 | −17 | −21 | 1.00E‐04 | 3.719017 | No Gray Matter found |
| 3 | 11 | −21 | −20 | 2.00E‐04 | 3.540084 | Right Brainstem.Midbrain.*.Gray Matter.Substania Nigra |
| 4 | −2.6 | 7.9 | 52.2 | 0 | 3.890594 | Left Cerebrum.Frontal Lobe.Medial Frontal Gyrus.Gray Matter.Brodmann area 6 |
| 5 | −30.6 | −8.8 | 59.1 | 0.00E+00 | 3.890594 | Left Cerebrum.Frontal Lobe.Precentral Gyrus.Gray Matter.Brodmann area 6 |
| 6 | 29.7 | 21.6 | 2.2 | 0 | 3.890594 | Right Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 7 | −43.2 | −35.3 | 45 | 0 | 3.890594 | Left Cerebrum.Parietal Lobe.Inferior Parietal Lobule.Gray Matter.Brodmann area 40 |
| 8 | 37 | 34.6 | 29.2 | 0 | 3.890594 | Right Cerebrum.Frontal Lobe.Middle Frontal Gyrus.Gray Matter.Brodmann area 9 |
| 9 | 29.2 | −4.7 | 49.9 | 0 | 3.890594 | Right Cerebrum.Frontal Lobe.Middle Frontal Gyrus.Gray Matter.Brodmann area 6 |
| 10 | 34.6 | −48.1 | 45 | 0 | 3.890594 | Right Cerebrum.Parietal Lobe.Superior Parietal Lobule.Gray Matter.Brodmann area 7 |
| 10 | 34.2 | −56.4 | 44.2 | 1.00E‐04 | 3.719017 | Right Cerebrum.Parietal Lobe.Angular Gyrus.Gray Matter.Brodmann area 39 |
| 11 | −45.3 | 0.9 | 35.5 | 0 | 3.890594 | Left Cerebrum.Frontal Lobe.Precentral Gyrus.Gray Matter.Brodmann area 6 |
| 12 | 44.8 | −37.5 | 44.8 | 0 | 3.890594 | Right Cerebrum.Parietal Lobe.Inferior Parietal Lobule.Gray Matter.Brodmann area 40 |
| 12 | 46.3 | −29.7 | 45.4 | 1.00E‐04 | 3.719017 | Right Cerebrum.Parietal Lobe.Inferior Parietal Lobule.Gray Matter.Brodmann area 40 |
| 13 | −28 | 44 | 14 | 0.0039 | 2.660607 | Left Cerebrum.Frontal Lobe.Middle Frontal Gyrus.Gray Matter.Brodmann area 10 |
FIGURE 4.
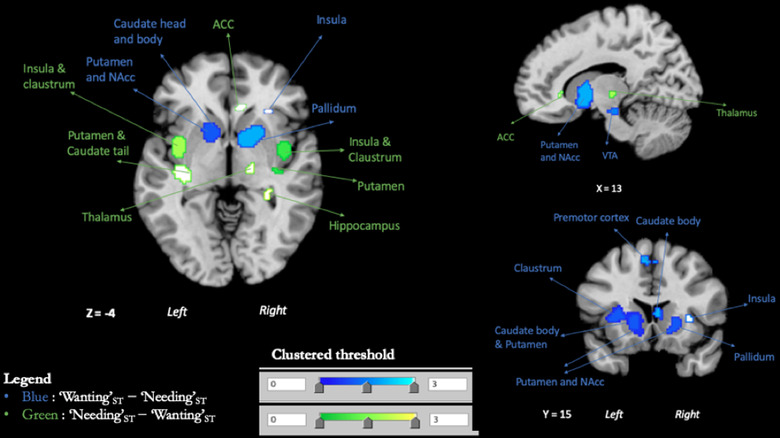
Contrasts maps. In blue, clustered thresholded maps for clusters of subtraction {[“Wanting”ST] minus [“Needing”ST]}. In green, clustered thresholded maps for clusters of subtraction {[“Needing”ST] minus [“Wanting”ST]}
3.3.3. Contrast: “Needing”ST–“Wanting”ST
Compared to “Wanting”ST, “Needing”ST more consistently activated regions of the right mid‐posterior insula, bilateral claustrum, left putamen (encompassing the tail of caudate), right anterior cingulate area, right thalamus, and bilateral hippocampus (Table 8 and Figure 4).
TABLE 8.
Coordinates for peak activated clusters in the Contrast: ‘Wanting’ST–‘Needing’ST
| Cluster # | x | y | z | P | Z | Label (nearest gray matter within 5 mm) |
|---|---|---|---|---|---|---|
| 1 | 44.9 | −6.9 | 1.5 | 0.00E+00 | 3.890594 | Right Cerebrum.Sub‐lobar.Insula.Gray Matter.Brodmann area 13 |
| 1 | 41.5 | −8.5 | −2 | 1.00E‐04 | 3.719017 | Right Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 1 | 38.3 | −14.6 | 5.1 | 2.00E‐04 | 3.540084 | Right Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 1 | 40 | −3 | −11 | 5.00E‐04 | 3.290527 | Right Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 2 | −40.3 | −7 | −2.7 | 2.00E‐04 | 3.540084 | Left Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 2 | −37.4 | −2.3 | −2.9 | 3.00E‐04 | 3.431614 | Left Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 3 | −30.9 | −24.6 | −0.6 | 1.00E‐04 | 3.719017 | Left Cerebrum.Sub‐lobar.Lentiform Nucleus.Gray Matter.Putamen |
| 3 | −37.5 | −20.2 | −2.4 | 5.00E‐04 | 3.290527 | Left Cerebrum.Sub‐lobar.Claustrum.Gray Matter.* |
| 3 | −34 | −20 | −10 | 2.00E‐03 | 2.878162 | Left Cerebrum.Temporal Lobe.Sub‐Gyral.Gray Matter.Hippocampus |
| 4 | 17 | −20 | −2 | 2.60E‐03 | 2.794376 | Right Cerebrum.Sub‐lobar.Thalamus.Gray Matter.Ventral Posterior Medial Nucleus |
| 5 | 28 | −36 | −6 | 3.00E‐03 | 2.747781 | Right Cerebrum.Temporal Lobe.Sub‐Gyral.Gray Matter.Hippocampus |
| 5 | 28 | −40 | 0 | 0.0082 | 2.39989 | Right Cerebrum.Temporal Lobe.Sub‐Gyral.Gray Matter.Hippocampus |
| 5 | 29.3 | −40 | −4.7 | 0.0083 | 2.39545 | Right Cerebrum.Temporal Lobe.Sub‐Gyral.Gray Matter.Hippocampus |
| 6 | 8 | 30 | −4 | 0.0029 | 2.758879 | Right Cerebrum.Limbic Lobe.Anterior Cingulate.Gray Matter.* |
4. DISCUSSION
Our goal was to compare the brain activation patterns related to value that comes from the state of “Wanting”ST from the one from the state of “Needing”ST. Our study was thus about when needing a stimulus influences its processing without the wanting component for that stimulus, and when “Wanting”ST happens without “Needing”ST. To answer this, we used an ALE neuroimaging meta‐analysis, comparing consistent brain activation patterns during processing of stimuli in these two states. We used the perception of a cue predicting a reward for “Wanting”ST, and we used the perception of food stimuli in a hungry state as a model for “Needing”ST. We first carried out separate meta‐analyses on “Wanting”ST and on “Needing”ST, then we contrasted and intersected them to identify differences and similarities between each of these states. We show that processing a stimulus in a “Wanting”ST state seems more related to activity within the mesolimbic dopaminergic brain areas, nigrostriatal dopaminergic regions, and striatal regions, while processing a stimulus in a “Needing”ST state seems more related to activity in viscerosensory cortices (e.g., mid‐posterior insula) and caudal‐ventral putamen (and to some extent the caudate tail). Both states seemed to share consistent activation in the caudate nucleus (head and body) and anterior insula. Compared to “Needing”ST, “Wanting”ST more consistently activated the mesolimbic dopamine: the VTA and ventral striatum and pallidum, and nigrostriatal dopamine regions (i.e., SN and dorsal striatum). Compared to “Wanting”ST, “Needing”ST more consistently activated the mid‐posterior insula and ACC, caudo‐ventral putamen and Caudate tail, and hippocampus. In the following paragraphs, we will discuss our overall results (the ones consistently found in all our meta‐analyses) and how by identifying the brain areas most implicated for each state (“Wanting”ST vs. “Needing”ST) can help us understand how we attribute different types of value to stimuli.
4.1. Overview of consistent activation patterns for Wanting versus Needing
Overall, our results—from main individual meta‐analyses, LOEO analyses, and contrasts—confirm that the activation pattern of “Wanting”ST related value shows consistent activation of VTA, ventral striatum, putamen, pallidum, and anterior insula. Our results are in line with previous human studies using a wide range of methods or approaches (Carter et al., 2009; Knutson et al., 2001, 2003; Krebs et al., 2009; O'Doherty, 2004; O'Doherty et al., 2002; Oldham et al., 2018; Schott et al., 2008; Simon et al., 2015; Wilson et al., 2018). For “Needing”ST, our results show that only the middle insula and to some extent the caudal‐ventral putamen are consistently related to “Needing”ST related value. The implication of the insula and dorsal striatum in “Needing”ST is in accordance with some previous literature findings (Goldstone et al., 2009; Siep et al., 2009; van der Laan et al., 2011). However, previous meta‐analyses and studies on “Needing”ST had also identified other regions such as OFC, ACC, and amygdala/parahippocampal gyrus (Chen & Zeffiro, 2020; Führer et al., 2008; LaBar et al., 2001; Mohanty et al., 2008). This could be due to the fact that we report here, regions that have been consistently found in all our meta‐analyses (main, contrasts, and LOEO) and thus use a more stringent approach than in previous meta‐analyses. Indeed, when only looking at results from our main meta‐analysis, we also identified regions within the OFC, ACC, and the amygdala. Nevertheless, using a more stringent approach, our results showing consistent activation mainly restricted to the mid‐posterior insula make sense as it is often considered as the core viscerosensory cortex because it projects to other visceromotor cortices (anterior insula, OFC, ACC) (Barrett & Simmons, 2015); and dense multimodal sensory interoceptive prediction errors converge within the posterior insula to guide interoception (Gehrlach et al., 2020). Thus, by combining contrasts, individual and LOEO meta‐analyses approaches, we were able to show that the core regions for “Needing”ST (in this case hunger) seems to be the mid‐posterior insula).
4.2. “Wanting”ST is more of an emotion than “Needing”ST
Our conjunction results showing consistent activations within the anterior insula for both “Wanting”ST and “Needing”ST could be related to the fact that this region integrates emotional states and is associated with emotional representation of internal states (Craig, 2010), and increases the significance of external stimuli that are relevant with regard to bodily, affective, and sensory information (Menon & Uddin, 2010; Young & Nusslock, 2016). Moreover, based on the fact that the anterior insula plays an important role in awareness (Craig, 2011), our findings suggest that this common activation could be related to our ability to be aware of our wants and needs. It is important to note that the anterior insula was found in the contrast “Wanting”ST–“Needing”ST, but not in “Needing”ST–“Wanting”ST. “Wanting”ST, viewed as reward seeking, is often considered as an emotional state (Panksepp, 2004) that may recruit the anterior insula during reward anticipation without necessity of “Needing”ST (Knutson et al., 2001; see Craig, 2010). Thus, “Wanting”ST can be thought of as a form of emotional reaction triggered by a Pavlovian cue that predicts a reward. Whereas “Needing”ST (physiological in this case) is usually considered a homeostatic emotion or sometimes a simple sensation, because physiological needs such as hunger do not seem to meet the criteria to be classified as emotions (see Panksepp, 2004), even though they can be seen as homeostatic emotions (Craig, 2003). In light of this, we speculate that, in humans, “Wanting”ST can have more emotional power than “Needing”ST because of the more consistent recruitment of the anterior insula. Though the anterior insula might contribute to turning “Wanting”ST into a more conscious desire/craving (Garavan, 2010; Naqvi et al., 2014), “Wanting”ST can also influence behavior without explicit awareness (Berridge & Robinson, 2003; Strack & Deutsch, 2004; Wei et al., 2017).
4.3. Short‐term value for “Wanting”ST versus long‐term value for “Needing”ST
While consistent activations were found for both states within the striatum, each seems to recruit a different sub‐region with the ventral and rostral parts, that is, NAcc, ventromedial caudate and rostroventral putamen more consistenly found for “Wanting”ST and the caudo‐ventral part of the putamen (that often included the tail of the caudate) more consistently found for “Needing”ST. This spatial difference could be related to the functional roles of these subregions including the coding of short‐ versus long‐term values of stimuli. Indeed, our results for “Wanting”ST are in line with findings that suggest that ventral striatum is more responsive to reward or its prediction than the dorsal striatum (Schultz et al., 2000) and that rostral striatum, mainly the caudate head, encodes short term or flexible value (Kim & Hikosaka, 2015). This is also in line with the view of “Wanting”ST as a moment to moment modulation of a cue that predicts reward in synergy with dopaminergic states (Zhang et al., 2009). In contrast, “Needing”ST, was more associated with consistent activation within the caudo‐ventral putamen (called “putamen tail”, see Kunimatsu et al., 2019) and (to a lesser extent) the caudate tail, both referred to as striatum tail (Amita et al., 2019), regions that acquire long‐term values of stimuli based on the historical experience of reward, but not on prediction of rewards (Kunimatsu et al., 2019). Thus, in line with theories and previous studies (Amita et al., 2019; Kim & Hikosaka, 2015; Kunimatsu et al., 2019; Zhang et al., 2009), our results might be interpreted as showing that value representation of a wanted versus needed stimuli rely on distinct regions of the striatum and that this difference could be driven by the temporal aspects or requirement of value processing for each state.
4.4. Directional and activational effect of value
The value assigned to stimuli can have a directional effect or activational effect. The directional effect is linked to choice (preference or action selection) and directs towards or away from stimuli, while the activational effect is related to action initiation, maintenance, and vigor (see Salamone et al., 2018). “Wanting”ST AND “Needing”ST meta‐analytic conjunction showed that both states consistently activate the caudate nucleus (head and body) and anterior insula (discussed above), regions implicated in action selection (Hollon et al., 2014; Ito & Doya, 2015; Petzschner et al., 2021) and emotional representation of internal states (Craig, 2010). The caudate is involved in goal‐directed behavior (Balleine & O'Doherty, 2010; Knutson & Cooper, 2005), and in the pairing between an action and the value of its consequence (Schwabe & Wolf, 2010), such as on the current state of the organism (see Balleine, 1992). Thus, the caudate is implicated in choice/action selection‐related value (Hollon et al., 2014; Ito & Doya, 2015), and is involved in directional value (Salamone et al., 2016). The implication of the caudate in “Wanting”ST AND “Needing”ST conjunction suggests both states can influence choice/action selection, that is, directional value of stimuli. Thus, by doing a meta‐analytic conjunction of “Wanting”ST AND “Needing”ST, we were able to show that both “Wanting”ST and “Needing”ST can influence the directional value of stimuli. However, as we will see, each state seems to rely on distinct neural substrates to compute this directional value.
Directional value for “Wanting”ST seems to arise from activity within the dopaminergic system. The VTA and SN, which contain the main dopaminergic neurons, were shown to be more consistently activated for “Wanting”ST than “Needing”ST. Our results also show that the regions of the ventral striatum, that is, the NAcc and the ventromedial caudate and rostroventral putamen (Haber & Knutson, 2010) were more consistently activated for “Wanting”ST–“Needing”ST, as well as the globus pallidus and the ventral pallidum (VP) (not shown). Indeed, incentive salience “Wanting”ST is generated when a reward cue is synergistically mixed with the state of mesocorticolimbic circuits (which mainly implicates the VTA, NAcc, and pallidum) (Warlow & Berridge, 2021; Zhang et al., 2009). Based on our results, we suggest that the directional value of “Wanting”ST towards stimuli comes from the cortico‐striato‐midbrain pathway, and first starts with the VTA which computes the prediction error that signals change in expected reward prediction (Schultz et al., 1997) and project mesolimbic dopamine to the ventral striatum (NAcc and VP) (Haber & Knutson, 2010). Second, the activity of the NAcc shell which corresponds to ventrolateral putamen in humans is the final path to the directional value of “Wanting”ST (Holmes et al., 2010), and it is known that mesolimbic dopamine activation within the NAcc or ventral striatum has strong influence on the dorsal striatum (Tricomi et al., 2009).
As mentioned before, with regard to “Needing”ST (the reaction to a needed stimulus), the middle insula, which was found as peak in all of our analyses including our contrasts in favour of “Needing”ST, seems to be the core regions for “Needing”ST (or in this case hunger: when one perceives food while hungry). In this sense, our results confirm that within the insula, it is the middle insula that pairs internal states to relevant external stimuli as argued by Craig (2010). Moreover, our findings dovetail those in the literature that show that the insula plays a role in an “as–if” representation of the bodily state (Damasio, 1994; Naqvi & Bechara, 2010), and that the insula encodes the incentive value of outcomes as a form of incentive memory (Balleine & Dickinson, 2000). Indeed, when hungry or thirsty, the mid‐posterior insula simulates future satiety state in the presence of food or water cues for both humans and animals (Chen et al., 2016; Livneh et al., 2020). Those cues create an interoceptive “prediction error” (see Barrett & Simmons, 2015). Based on interoceptive prediction error from mid‐posterior insula, the visceromotor cortices (ACC, OFC, and anterior insula) make predictions about desired internal states (Barrett & Simmons, 2015), and enhance the value of stimuli and actions that fulfill the predictions (Petzschner et al., 2021). Based on our results, we suggest that the mid‐posterior insula prediction error might be the origin of the directional value of “Needing”ST in the same logic the VTA does for “Wanting”ST, that is, by computing a sort of prediction error that influences cue selection (see Arsenault et al., 2014); and in this case (i.e., for “Needing”ST), it is an interoceptive prediction error (Barrett & Simmons, 2015). In this regard, the directional value of “Wanting”ST and that of “Needing”ST depends on two different prediction errors: for “Wanting”ST, the prediction error is computed within the VTA, and for “Needing”ST, the (interoceptive) prediction error is computed within the mid‐posterior insula. Importantly, although we focused our meta‐analysis on the hunger state and the processing of food stimuli, we think that our results can be generalized to other types of needing states and stimuli. Indeed, it is known that the mid‐posterior insula receives multimodal sensory interoceptive signals to compute a prediction error (Livneh et al., 2020).
If both states can give rise to directional/action selection value (albeit differently), only “Wanting”ST seems associated to activiational value. Indeed, consistent activations within the NAcc was only found in our “Wanting”ST meta‐analysis, “Wanting”ST–“Needing”ST meta‐analytic contrast, and even when stringent LOEO analyses were used. “Wanting”ST has more (compared to “Needing”ST) control on activational value because the prediction error signal is sent to the NAcc, which (makes those predictions and) has strong influence to the pallidum which has a lot of impact on invigoration of motor action possibly through a more direct connection to the thalamus (Balleine & O'Doherty, 2010; Haber & Knutson, 2010). In line with literature, our results point out that the activational aspects of cue‐induced “Wanting”ST is likely mediated by the mesolimbic dopamine that implicates activation within the central NAcc (see Holmes et al., 2010; Salamone & Correa, 2002; Salamone et al., 1997, 2018, 2016) or ventral striatal regions in general (Haber & Knutson, 2010), which have strong influence on the dorsal striatum (Tricomi et al., 2009). Our results that “Needing”ST did not consistently activate dopaminergic regions, are in line with the now admitted fact that needs by themselves do not have activational value (see Salamone et al., 2018) and are not the main source of motivated behavior (Bindra, 1974; Berridge, 2004), although they can amplify it (Toates, 1994). The fact that “Needing”ST has only the directional part (choice/preference or action selection), not the activational one, means that a needed stimulus must still become “wanted,” by altering mesolimbic dopamine reactivity and encountering a relevant reward predicting cue (Zhang et al., 2009), in order to have full motivational value (Berridge, 2004; Bindra, 1974; Toates, 1994). Thus, motivation is better explained by incentive salience “Wanting”ST than by “Needing”ST (Berridge, 2004; Bindra, 1974). Nevertheless, “Needing”ST can affect “liking” (see Berridge, 2009) (whether for hunger and food or thirst and water) (Balleine, 1992; Dayan & Balleine, 2002) and can create expectation of “liking” through “cognitive desire” (see Berridge, 2012) towards a needed stimulus. This latter is more goal oriented, and based on declarative memories and on cognitive expectations of act–outcome relations (Berridge, 2012). Thus, “Needing”ST generates cognitive desire, but not necessarily “Wanting”ST (incentive salience) (Berridge, 2012).
One can wonder, if “Needing”ST does not provide activational value to stimuli, what motivates exploratory behavior in the hungry state? We think that such exploratory behavior, in absence of any reward cue, is related to what has been called the “seeking system” by Panksepp (2004). This system is composed of the hypothalamus, the ventral striatum and the VTA (Panksepp, 2004), and can include the ventromedial prefrontal cortex (Di Domenico & Ryan, 2017; Panksepp & Biven, 2012), and is responsible for energized exploratory and search behaviors and investigation, and it does not need to be stimulated by a positive incentive cue (Harmon‐Jones et al., 2013; Panksepp, 2004). However, when there is a reward cue, activity of that system contributes to adding incentive salience or “Wanting”ST to that cue (Berridge, 2004) and thus turn the exploratory behavior into need‐induced “Wanting”ST (see Anselme, 2015). Moreover, it should also be noted that need states can direct and somehow elicit behavior even in absence of the energizing effect of incentive salience “Wanting”ST (Balleine, 2005; Niv et al., 2006; Salamone et al., 2018), and areas of the ventromedial prefrontal cortex and ACC can elicit goal directed behavior based on internal states, even in absence of a reward cue that prompts and guides the animal (Passingham & Wise, 2012).
4.5. Incentive cue (“Wanting”ST) versus outcome relevant cue (“Needing”ST)
Although “Wanting”ST and “Needing”ST are constructs that surely extend beyond the kind of reward, the interpretation of the present results should be done with caution as some brain areas might be indeed influenced by the type of reward, namely the difference between primary (food for “Needing”ST experiments) and secondary (money or points for “Wanting”ST experiments) reward. Of note, “Wanting”ST studies did not explicitly exclude food‐related studies and were not limited to secondary rewards, but our selection criteria resulted in the fact that we did not find food‐related “Wanting”ST studies to include. However, we believe that, in our study, the difference is not in terms of the kind of reward, but in terms of situations or paradigms, as was also argued by Sescousse et al. (2013) who conducted a meta‐analysis between primary and secondary reward, and found higher dopaminergic striatal activation for secondary reward. They argued that it “is unlikely to be related to the very nature of monetary rewards,” but more about the “protocols used” (Sescousse et al., 2013). In other words, it is unlikely that there is more dopaminergic activation within the ventral striatum for monetary (secondary) reward compared to food or sex (primary) reward, and the difference is indeed in the paradigm (situation) used rather than the type of reward (Sescousse et al., 2013). In our view, the cues in the “Wanting”ST experiments were incentive stimuli, that is, stimuli that motivate action and bias behavior (see Robinson et al., 2014); whereas the reward cues included in the “Needing”ST experiments were not, though they were relevant for the current (deprivational) state (see Balleine, 2005). Thus, though the type of reward can influence some brain activation, it is the role (rather than nature) of the reward cue, based on paradigms or situation, that mainly differentiate between “Wanting”ST and “Needing”ST, and generate either mesolimbic‐related reward prediction/prediction error or interoceptive prediction/prediction error, respectively. In a similar paradigm (i.e., where a food cue triggers action to gain it), hunger and/or food (used here for “Needing”ST) would also activate “Wanting”ST, and would result in dopamine‐related ventral striatum activation (Simon et al., 2016, 2015; Yousuf et al., 2018). Of note, “Wanting”ST studies did not explicitly exclude food‐related studies and were not limited to secondary rewards. In “Wanting”ST experiments that include food, food cues elicited higher mesolimbic dopaminergic response in the ventral striatum when participants were hungry, and such activity was reduced for food cues when participants were satiated (Yousuf et al., 2018). This can be viewed as hunger elevating mesolimbic dopamine, which peaked at the event of the food cue as argued in incentive salience theory (Berridge, 1996, 2004). Thus, “Wanting”ST is closely related to the presence of a cue, associated to either primary or secondary reward, that triggers action or approach behavior based on mesolimbic dopamine state, and that can happen with or without need state (see DiFeliceantonio & Berridge, 2012). Though we found no study in which participants are in need of money, it is possible that passively viewing money pictures would not necessarily lead to significant mesolimbic dopamine activity, that is, “Wanting”ST, if those pictures are not cues to pursue monetary rewards. Also, it is not sure whether, if one needs money, passively viewing money cues would trigger activity in the mid‐posterior insula in the same way as food/hunger or water/thirst (Livneh et al., 2020). Both these hypotheses would need to be empirically tested. In brief, “Wanting”ST and “Needing”ST provide two different roles/values to reward cues, depending on situations or paradigms. “Wanting”ST related cues are stimuli that motivate action and bias behavior (see Robinson et al., 2014), whereas the “Needing”ST related cues are stimuli that are outcome relevant for the current need state (see Balleine, 2005), and those two roles/values do not necessarily (or always) go together. In that sense, our findings and discussion are more about a difference between such cue roles/values, rather than a difference between primary and secondary reward.
4.6. Implication on addiction and other maladaptive behaviors
The conceptualization of “Needing”ST, when it happens without “Wanting”ST somehow dovetails recent neurobehavioral theories of addiction (including food addiction, but see Gearhardt et al., 2011) (for a review see Bickel et al., 2018). Our study, showing VTA and ventral striatum for “Wanting”ST, mid‐posterior insula for “Needing”ST, and anterior insula for both, allow us to speculate that there might be at least two sources of origin of addiction‐like behavior. One would be related to interoception, likely gated by the insula, with the mid‐posterior insula receiving aversive states information (Gehrlach et al., 2020; Livneh et al., 2020) as found in our “Needing”ST–“Wanting”ST contrast; and anterior insula turning such information into emotion, desire and craving (Craig, 2009; Noel et al., 2013; Turel & Bechara, 2016) as shown in our “Wanting”ST–“Needing”ST contrast and conjunction map. Whereas the other source of addictive behavior would be related to dopamine activity, mainly in the striatum (Berridge & Robinson, 2016; Hogarth et al., 2013; Robinson & Berridge, 1993; Volkow et al., 1999) as found in our “Wanting”ST–“Needing”ST contrast. This could lead to difference between “addiction” and “dependence” (see O'Brien et al., 2006), with addiction being related to excessive consumption, and thus more related to “Wanting”ST. Whereas dependence would be more related to aversive states caused by withdrawal symptoms (that can happen in absence of addiction, see O'Brien et al., 2006), and thus more closer to “Needing”ST. The difference could also be temporal, with the beginning of addiction and establishment of sensitization related to striatal dopaminergic areas, thus “Wanting”ST; and withdrawal symptoms due to aversives states related to perception of aversive internal states and urge to terminate them within the insula, which includes “Needing”ST. Moreover, dopamine‐related sensitization theory often separates “liking” from “wanting,” where the latter is more responsible for addictive behavior (Berridge & Robinson, 2016). It might be possible to add “Needing”ST as a separate component. In that case “liking”ST can serve as the beginning of the addictive process, but soon then sensitization happens and is mediated by dopamine‐related areas and “Wanting”ST, and withdrawal is mediated by insula and “Needing”ST.
4.7. Limits
We would like to point out some limits to our work. First, “Wanting”ST experiments did not include physiologically related stimuli such as food or water. We argue that the experiments for “Wanting”ST really expressed “wanting” (see Berridge, 2004) because the contrasts we used focused on the processing of the cues, not the outcome, and the cues in those experiments triggered the decision for reward seeking. Indeed, “wanting” has been related to decision utility, that is, the “choice to pursue or consume an outcome” (Berridge & O'Doherty, 2014), induced by a cue (Berridge & Aldridge, 2009). So, although we could not know the mesolimbic state of participants in those experiments, the behavioral situations, that is, a cue that triggered a decision to seek reward, do seem to induce “wanting” (see Berridge, 2004). Furthermore, “Wanting”ST or motivation activates a general system, regardless of the type of stimulus (Bouton, 2016). Thus, though most stimuli for “Wanting”ST were money or points, the mechanism for cue induced decision for reward seeking, that is, “Wanting”ST is the same.
Second, “Needing”ST contrasts only focused on hunger and the processing of food (physiologically related stimuli). Though our method could seem dependent on the type of physiological state, that is, hunger, and thus the type of needed stimuli, that is, food; our findings and interpretation seem to go in the same direction than studies and theories that suggest an integrative role of all physiological states within the insula. In that sense, Gehrlach et al.’s (2020) results suggest that dense multimodal sensory prediction errors converge in the posterior insula to guide interoception. Based on that, it has been argued that that the insula represents physiological state in cue‐independent spontaneous activity, which is then modified directionally by cues that predict water or food availability (Namboodiri & Stuber, 2020). This suggests that our findings regarding the activation of mid‐posterior insula might not be hunger/food dependent but rather be resulting from a more general role of the insula with regard to bodily states, specifically serving interoceptive inference (Allen, 2020). Though there are justifications for our methodology in general, it would still be interesting for future studies to test the difference between “Wanting”ST and “Needing”ST with more control on the dopaminergic state, on the type of physiological states and on the type of stimuli.
Nevertheless, wanting and/or needing go way beyond eating, drinking or winning money or points. The distinction between wanting versus needing can apply to virtually any decision, and can even be viewed as philosophical or phenomenological concepts. In that sense, they go way beyond our study and the tools we have used. Thus, our study should be viewed as testing some manifestation of wanting and/or needing rather than testing the general phenomena.
5. CONCLUSION
Our goal was to compare the brain representation of “Wanting”ST and “Needing”ST related values —two states that guide value attribution and our consumption behaviors. Our results suggest distinct brain systems for both states, with the mesolimbic dopaminergic circuitry as the core for “Wanting”ST and the posterior‐middle insula for “Needing”ST. Whereas “Needing”ST only provides directional value (through an interoceptive prediction error), “Wanting”ST which involves dopamine provides both directional (through a reward prediction error) and activational value. Because “Needing”ST does not provide activational value to stimuli, full motivation (directional and activational) to consume depends more on “Wanting”ST than on “Needing”ST, and means that “Wanting”ST has more power to activate behavior. This might explain why we consume what we want beyond what we need (Stern, 2000).
CONFLICT OF INTEREST
The authors declare that they have no conflict of interest.
AUTHOR CONTRIBUTIONS
Juvenal Bosulu: Designed the study, performed the database search, performed data analysis, interpretation, and wrote the manuscript. Sébastien Hétu: Designed the study, revised the manuscript and provided critical feedbacks. Max‐Antoine Allaire: Performed the database search, revised the manuscript and provided critical feedbacks. Laurence Tremblay‐Grenier: Performed the database search, revised the manuscript and provided critical feedbacks. Yi Luo: Revised the manuscript and provided critical feedbacks. Simon Eichkoff: Revised the manuscript and provided critical feedbacks. All authors contributed to and approved the final manuscript version.
PEER REVIEW
The peer review history for this article is available at https://publons.com/publon/10.1002/brb3.2713
Supporting information
Supplementary material
ACKNOWLEDGMENT
The research was supported in part by NSERC Discovery Grant #RGPIN‐2018‐05698, UdeM institutional funds, and Mitacs Grant #IT20458.
Bosulu, J. , Allaire, M.‐A. , Tremblay‐Grénier, L. , Luo, Y. , Eickhoff, S. , & Hétu, S. (2022). “Wanting” versus “needing” related value: An fMRI meta‐analysis. Brain and Behavior, 12, e2713. 10.1002/brb3.2713
DATA AVAILABILITY STATEMENT
All data are available upon request.
REFERENCES
- Allen, M. (2020). Unravelling the neurobiology of interoceptive inference. Trends in Cognitive Sciences, 24(4), 265–266. 10.1016/j.tics.2020.02.002 [DOI] [PubMed] [Google Scholar]
- Anselme, P. (2015). Incentive salience attribution under reward uncertainty: A Pavlovian model. Behavioural processes, 111, 6–18. 10.1016/j.beproc.2014.10.016 [DOI] [PubMed] [Google Scholar]
- Amita, H. , Kim, H. F. , Smith, M. K. , Gopal, A. , & Hikosaka, O. (2019). Neuronal connections of direct and indirect pathways for stable value memory in caudal basal ganglia. European Journal of Neuroscience, 49(5), 712–725. 10.1111/ejn.13936 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Appleton, K. M. , Gentry, R. C. , & Shepherd, R. (2006). Evidence of a role for conditioning in the development of liking for flavours in humans in everyday life. Physiology & Behavior, 87(3), 478–486. 10.1016/j.physbeh.2005.11.017 [DOI] [PubMed] [Google Scholar]
- Arsenault, J. T. , Rima, S. , Stemmann, H. , & Vanduffel, W. (2014). Role of the primate ventral tegmental area in reinforcement and motivation. Current Biology, 24(12), 1347–1353. 10.1016/j.cub.2014.04.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balleine, B. (1992). Instrumental performance following a shift in primary motivation depends on incentive learning. Journal of Experimental Psychology: Animal Behavior Processes, 18(3), 236. 10.1037/0097-7403.18.3.236 [DOI] [PubMed] [Google Scholar]
- Balleine, B. W. (2005). Neural bases of food‐seeking: Affect, arousal and reward in corticostriatolimbic circuits. Physiology & behavior, 86(5), 717–730. 10.1016/j.physbeh.2005.08.061 [DOI] [PubMed] [Google Scholar]
- Balleine, B. W. (2009). Taste, disgust, and value: Taste aversion learning and outcome encoding in instrumental conditioning. In: Reilly TRS, S. (Ed.), Conditioned taste aversion: Behavioral and neural processes (pp. 262–280). Oxford University Press. [Google Scholar]
- Balleine, B. W. , & Dickinson, A. (2000). The effect of lesions of the insular cortex on instrumental conditioning: Evidence for a role in incentive memory. Journal of Neuroscience, 20(23), 8954–8964. 10.1523/JNEUROSCI.20-23-08954.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balleine, B. W. , & Killcross, S. (2006). Parallel incentive processing: An integrated view of amygdala function. Trends in Neurosciences, 29(5), 272–279. 10.1016/j.tins.2006.03.002 [DOI] [PubMed] [Google Scholar]
- Balleine, B. W. , & O'Doherty, J. P. (2010). Human and rodent homologies in action control: Corticostriatal determinants of goal‐directed and habitual action. Neuropsychopharmacology, 35(1), 48–69. 10.1038/npp.2009.131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrett, L. F. , & Simmons, W. K. (2015). Interoceptive predictions in the brain. Nature Reviews Neuroscience, 16(7), 419–429. 10.1038/nrn3950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge, K. C. (1996). Food reward: Brain substrates of wanting and liking. Neuroscience & Biobehavioral Reviews, 20(1), 1–25. 10.1016/0149-7634(95)00033-B [DOI] [PubMed] [Google Scholar]
- Berridge, K. C. (2004). Motivation concepts in behavioral neuroscience. Physiology & Behavior, 81(2), 179–209. 10.1016/j.physbeh.2004.02.004 [DOI] [PubMed] [Google Scholar]
- Berridge, K. C. (2009). ‘Liking’ and ‘wanting’ food rewards: Brain substrates and roles in eating disorders. Physiology & Behavior, 97(5), 537–550. 10.1016/j.physbeh.2009.02.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge, K. C. (2012). From prediction error to incentive salience: Mesolimbic computation of reward motivation. European Journal of Neuroscience, 35(7), 1124–1143. 10.1111/j.1460-9568.2012.07990.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge, K. C. (2018). Evolving concepts of emotion and motivation. Frontiers in Psychology, 9, 1647. 10.3389/fpsyg.2018.01647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge, K. C. , & Aldridge, J. W. (2009). Decision utility, incentive salience, and cue‐triggered “wanting.”. Oxford series in Social Cognition and Social Neuroscience, 2009, 509. [PMC free article] [PubMed] [Google Scholar]
- Berridge, K. C. , & O'Doherty, J. P. (2014). From experienced utility to decision utility. In Neuroeconomics (pp. 335–351). Academic Press. 10.1016/B978-0-12-416008-8.00018-8 [DOI] [Google Scholar]
- Berridge, K. C. , & Robinson, T. E. (1998). What is the role of dopamine in reward: Hedonic impact, reward learning, or incentive salience? Brain research reviews, 28(3), 309–369. 10.1016/S0165-0173(98)00019-8 [DOI] [PubMed] [Google Scholar]
- Berridge, K. C. , & Robinson, T. E. (2003). Parsing reward. Trends in Neurosciences, 26(9), 507–513. 10.1016/S0166-2236(03)00233-9 [DOI] [PubMed] [Google Scholar]
- Berridge, K. C. , & Robinson, T. E. (2016). Liking, wanting, and the incentive‐sensitization theory of addiction. American Psychologist, 71(8), 670. 10.1037/amp0000059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bickel, W. K. , Mellis, A. M. , Snider, S. E. , Athamneh, L. N. , Stein, J. S. , & Pope, D. A. (2018). 21st century neurobehavioral theories of decision making in addiction: Review and evaluation. Pharmacology Biochemistry and Behavior, 164, 4–21. 10.1016/j.pbb.2017.09.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bindra, D. (1968). Neuropsychological interpretation of the effects of drive and incentive‐motivation on general activity and instrumental behavior. Psychological Review, 75, 1–22. 10.1037/h0025306 [DOI] [Google Scholar]
- Bindra, D. (1974). A motivational view of learning, performance, and behavior modification. Psychological Review, 81(3), 199. 10.1037/h0036330 [DOI] [PubMed] [Google Scholar]
- Bolles, R. C. , & Moot, S. A. (1972). Derived motives. Annual Review of Psychology, 23(1), 51–72. 10.1146/annurev.ps.23.020172.000411 [DOI] [Google Scholar]
- Bouton, M. E. (2016). Learning and behavior: A contemporary synthesis. Sinauer Associates. [Google Scholar]
- Brunstrom, J. M. , & Fletcher, H. Z. (2008). Flavour–flavour learning occurs automatically and only in hungry participants. Physiology & behavior, 93(1–2), 13–19. 10.1016/j.physbeh.2007.07.009 [DOI] [PubMed] [Google Scholar]
- Campbell, C. (1998). Consumption and the rhetorics of need and want. Journal of Design History, 11(3), 235–246. 10.1093/jdh/11.3.235 [DOI] [Google Scholar]
- Cardinal, R. N. , Parkinson, J. A. , Hall, J. , & Everitt, B. J. (2002). Emotion and motivation: The role of the amygdala, ventral striatum, and prefrontal cortex. Neuroscience & Biobehavioral Reviews, 26(3), 321–352. 10.1016/S0149-7634(02)00007-6 [DOI] [PubMed] [Google Scholar]
- Carter, R. M. , MacInnes, J. J. , Huettel, S. A. , & Adcock, R. A. (2009). Activation in the VTA and nucleus accumbens increases in anticipation of both gains and losses. Frontiers in Behavioral Neuroscience, 3, 21. 10.3389/neuro.08.021.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chein, J. M. , Fissell, K. , Jacobs, S. , & Fiez, J. A. (2002). Functional heterogeneity within Broca's area during verbal working memory. Physiology & Behavior, 77(4–5), 635–639. 10.1016/S0031-9384(02)00899-5 [DOI] [PubMed] [Google Scholar]
- Chen, E. Y. , & Zeffiro, T. A. (2020). Hunger and BMI modulate neural responses to sweet stimuli: FMRI meta‐analysis. International Journal of Obesity, 44(8), 1636–1652. 10.1038/s41366-020-0608-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, J. , Papies, E. K. , & Barsalou, L. W. (2016). A core eating network and its modulations underlie diverse eating phenomena. Brain and Cognition, 110, 20–42. 10.1016/j.bandc.2016.04.004 [DOI] [PubMed] [Google Scholar]
- Craig, A. D. (2003). Interoception: The sense of the physiological condition of the body. Current Opinion in Neurobiology, 13(4), 500–505. 10.1016/S0959-4388(03)00090-4 [DOI] [PubMed] [Google Scholar]
- Craig, A. D. (2009). How do you feel—Now? The anterior insula and human awareness. Nature Reviews Neuroscience, 10(1), 59–70. 10.1038/nrn2555 [DOI] [PubMed] [Google Scholar]
- Craig, A. D. (2010). The sentient self. Brain Structure and Function, 214, 563–577. 10.1007/s00429-010-0248-y [DOI] [PubMed] [Google Scholar]
- Craig, A. D. (2011). Significance of the insula for the evolution of human awareness of feelings from the body. Annals of the New York Academy of Sciences, 1225(1), 72–82. 10.1111/j.1749-6632.2011.05990.x [DOI] [PubMed] [Google Scholar]
- Damasio, A. R. (1994) Descartes’ Error: Emotion, reason and the human brain. Putnam and Sons. [Google Scholar]
- Dayan, P. , & Balleine, B. W. (2002). Reward, motivation, and reinforcement learning. Neuron, 36(2), 285–298. 10.1016/S0896-6273(02)00963-7 [DOI] [PubMed] [Google Scholar]
- Dickinson, A. , & Balleine, B. (1994). Motivational control of goal‐directed action. Animal Learning & Behavior, 22(1), 1–18. 10.3758/BF03199951 [DOI] [Google Scholar]
- Di Domenico, S. I. , & Ryan, R. M. (2017). The emerging neuroscience of intrinsic motivation: A new frontier in self‐determination research. Frontiers in Human Neuroscience, 11, 145. 10.3389/fnhum.2017.00145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiFeliceantonio, A. G. , & Berridge, K. C. (2012). Which cue to ‘want’? Opioid stimulation of central amygdala makes goal‐trackers show stronger goal‐tracking, just as sign‐trackers show stronger sign‐tracking. Behavioural Brain Research, 230(2), 399–408. 10.1016/j.bbr.2012.02.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff, S. B. , Bzdok, D. , Laird, A. R. , Kurth, F. , & Fox, P. T. (2012). Activation likelihood estimation meta‐analysis revisited. Neuroimage, 59(3), 2349–2361. 10.1016/j.neuroimage.2011.09.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff, S. B. , Bzdok, D. , Laird, A. R. , Roski, C. , Caspers, S. , Zilles, K. , & Fox, P. T. (2011). Co‐activation patterns distinguish cortical modules, their connectivity and functional differentiation. Neuroimage, 57(3), 938–949. 10.1016/j.neuroimage.2011.05.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff, S. B. , Laird, A. R. , Grefkes, C. , Wang, L. E. , Zilles, K. , & Fox, P. T. (2009). Coordinate‐based ALE meta‐analysis of neuroimaging data: A random‐effects approach based on empirical estimates of spatial uncertainty. Human Brain Mapping, 30(9), 2907. 10.1002/hbm.20718 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eklund, A. , Nichols, T. E. , & Knutsson, H. (2016). Cluster failure: Why fMRI inferences for spatial extent have inflated false‐positive rates. Proceedings of the National Academy of Sciences, 113(28), 7900–7905. 10.1073/pnas.1602413113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischl, B. , Stevens, A. A. , Rajendran, N. , Yeo, B. T. , Greve, D. N. , Van Leemput, K. , Polimeni, J. R. , Kakunoori, S. , Buckner, R. L. , Pacheco, J. , Salat, D. H. , Melcher, J. , Frosch, M. P. , Hyman, B. T. , Grant, P. E. , Rosen, B. R. , van der Kouwe, A. J. W. , Wiggins, G. C. , Wald, L. L. , & Augustinack, J. C. (2009). Predicting the location of entorhinal cortex from MRI. Neuroimage, 47(1), 8–17. 10.1016/j.neuroimage.2009.04.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Führer, D. , Zysset, S. , & Stumvoll, M. (2008). Brain activity in hunger and satiety: An exploratory visually stimulated FMRI study. Obesity, 16(5), 945–950. [DOI] [PubMed] [Google Scholar]
- Garavan, H. (2010). Insula and drug cravings. Brain Structure and Function, 214(5–6), 593–601. 10.1038/oby.2008.33 [DOI] [PubMed] [Google Scholar]
- Gearhardt, A. , A White, M. , & N Potenza, M. (2011). Binge eating disorder and food addiction. Current Drug Abuse Reviews, 4(3), 201–207. 10.2174/1874473711104030201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gehrlach, D. A. , Dolensek, N. , Klein, A. S. , Chowdhury, R. R. , Matthys, A. , Junghänel, M. , Gaitanos, T. N. , Podgornik, A. , Black, T. D. , Vaka, N. R. , Conzelmann, K.‐K. , & Gogolla, N. (2020). Aversive state processing in the posterior insular cortex. Nature Neuroscience, 22(9), 1424–1437. 10.1038/s41593-019-0469-1 [DOI] [PubMed] [Google Scholar]
- Gibson, E. L. , Wainwright, C. J. , & Booth, D. A. (1995). Disguised protein in lunch after low‐protein breakfast conditions food‐flavor preferences dependent on recent lack of protein intake. Physiology & Behavior, 58(2), 363–371. 10.1016/0031-9384(95)00068-T [DOI] [PubMed] [Google Scholar]
- Goldstone, A. P. , Prechtl de Hernandez, C. G. , Beaver, J. D. , Muhammed, K. , Croese, C. , Bell, G. , Durighel, G. , Hughes, E. , Waldman, A. D. , Frost, G. , & Bell, J. D. (2009). Fasting biases brain reward systems towards high‐calorie foods. European Journal of Neuroscience, 30(8), 1625–1635. 10.1111/j.1460-9568.2009.06949.x [DOI] [PubMed] [Google Scholar]
- Haber, S. N. , & Knutson, B. (2010). The reward circuit: Linking primate anatomy and human imaging. Neuropsychopharmacology, 35(1), 4–26. 10.1038/npp.2009.129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamid, A. A. , Pettibone, J. R. , Mabrouk, O. S. , Hetrick, V. L. , Schmidt, R. , Vander Weele, C. M. , Kennedy, R. T. , Aragona, B. J. , & Berke, J. D. (2016). Mesolimbic dopamine signals the value of work. Nature Neuroscience, 19(1), 117–126. 10.1038/nn.4173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harmon‐Jones, E. , Harmon‐Jones, C. , & Price, T. F. (2013). What is approach motivation? Emotion Review, 5(3), 291–295. 10.1177/1754073913477509 [DOI] [Google Scholar]
- Hogarth, L. , Balleine, B. W. , Corbit, L. H. , & Killcross, S. (2013). Associative learning mechanisms underpinning the transition from recreational drug use to addiction. Annals of the New York Academy of Sciences, 1282(1), 12–24. 10.1111/j.1749-6632.2012.06768.x [DOI] [PubMed] [Google Scholar]
- Hogarth, L. , & Chase, H. W. (2011). Parallel goal‐directed and habitual control of human drug‐seeking: Implications for dependence vulnerability. Journal of Experimental Psychology: Animal Behavior Processes, 37(3), 261. 10.1037/a0022913 [DOI] [PubMed] [Google Scholar]
- Hollon, N. G. , Arnold, M. M. , Gan, J. O. , Walton, M. E. , & Phillips, P. E. (2014). Dopamine‐associated cached values are not sufficient as the basis for action selection. Proceedings of the National Academy of Sciences, 111(51), 18357–18362. 10.1073/pnas.1419770111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes, N. M. , Marchand, A. R. , & Coutureau, E. (2010). Pavlovian to instrumental transfer: A neurobehavioural perspective. Neuroscience & Biobehavioral Reviews, 34(8), 1277–1295. 10.1016/j.neubiorev.2010.03.007 [DOI] [PubMed] [Google Scholar]
- Hull, C. L. (1943). Principles of behavior: An introduction to behavior theory. Appleton‐Century. [Google Scholar]
- Ikemoto, S. , & Panksepp, J. (1999). The role of nucleus accumbens dopamine in motivated behavior: A unifying interpretation with special reference to reward‐seeking. Brain Research Reviews, 31(1), 6–41. 10.1016/S0165-0173(99)00023-5 [DOI] [PubMed] [Google Scholar]
- Ito, M. , & Doya, K. (2015). Distinct neural representation in the dorsolateral, dorsomedial, and ventral parts of the striatum during fixed‐and free‐choice tasks. Journal of Neuroscience, 35(8), 3499–3514. 10.1523/JNEUROSCI.1962-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kern, D. L. , McPhee, L. , Fisher, J. , Johnson, S. , & Birch, L. L. (1993). The postingestive consequences of fat condition preferences for flavors associated with high dietary fat. Physiology & Behavior, 54(1), 71–76. 10.1016/0031-9384(93)90045-H [DOI] [PubMed] [Google Scholar]
- Kim, H. F. , & Hikosaka, O. (2015). Parallel basal ganglia circuits for voluntary and automatic behaviour to reach rewards. Brain, 138(7), 1776–1800. 10.1093/brain/awv134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knutson, B. , & Cooper, J. C. (2005). Functional magnetic resonance imaging of reward prediction. Current Opinion in Neurology, 18(4), 411–417. 10.1097/01.wco.0000173463.24758.f6 [DOI] [PubMed] [Google Scholar]
- Knutson, B. , Fong, G. W. , Adams, C. M. , Varner, J. L. , & Hommer, D. (2001). Dissociation of reward anticipation and outcome with event‐related fMRI. Neuroreport, 12(17), 3683–3687. [DOI] [PubMed] [Google Scholar]
- Knutson, B. , Fong, G. W. , Bennett, S. M. , Adams, C. M. , & Hommer, D. (2003). A region of mesial prefrontal cortex tracks monetarily rewarding outcomes: Characterization with rapid event‐related fMRI. Neuroimage, 18(2), 263–272. 10.1016/S1053-8119(02)00057-5 [DOI] [PubMed] [Google Scholar]
- Knutson, B. , Westdorp, A. , Kaiser, E. , & Hommer, D. (2000). FMRI visualization of brain activity during a monetary incentive delay task. Neuroimage, 12(1), 20–27. 10.1006/nimg.2000.0593 [DOI] [PubMed] [Google Scholar]
- Krebs, R. M. , Schott, B. H. , & Düzel, E. (2009). Personality traits are differentially associated with patterns of reward and novelty processing in the human substantia nigra/ventral tegmental area. Biological Psychiatry, 65(2), 103–110. 10.1016/j.biopsych.2008.08.019 [DOI] [PubMed] [Google Scholar]
- Kunimatsu, J. , Maeda, K. , & Hikosaka, O. (2019). The caudal part of putamen represents the historical object value information. Journal of Neuroscience, 39(9), 1709–1719. 10.1523/JNEUROSCI.2534-18.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaBar, K. S. , Gitelman, D. R. , Parrish, T. B. , Kim, Y. H. , Nobre, A. C. , & Mesulam, M. (2001). Hunger selectively modulates corticolimbic activation to food stimuli in humans. Behavioral Neuroscience, 115(2), 493. 10.1037/0735-7044.115.2.493 [DOI] [PubMed] [Google Scholar]
- Lex, A. , & Hauber, W. (2008). Dopamine D1 and D2 receptors in the nucleus accumbens core and shell mediate Pavlovian‐instrumental transfer. Learning & Memory, 15(7), 483–491. 10.1101/lm.978708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipschutz, R. D. (2001). Globalised networks of knowledge and practice: Civil society and environmental governance. In Global environmental policies (pp. 252–280). Palgrave Macmillan. 10.1057/9780230503359_9 [DOI] [Google Scholar]
- Livneh, Y. , Sugden, A. U. , Madara, J. C. , Essner, R. A. , Flores, V. I. , Sugden, L. A. , Resch, J. M. , Lowell, B. B. , & Andermann, M. L. (2020). Estimation of current and future physiological states in insular cortex. Neuron, 105(6), 1094–1111. 10.1016/j.neuron.2019.12.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menon, V. , & Uddin, L. Q. (2010). Saliency, switching, attention and control: A network model of insula function. Brain Structure and Function, 214(5–6), 655–667. 10.1007/s00429-010-0262-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Millman, Z. B. , Gallagher, K. , Demro, C. , Schiffman, J. , Reeves, G. M. , Gold, J. M. , & Waltz, J. A. (2020). Evidence of reward system dysfunction in youth at clinical high‐risk for psychosis from two event‐related fMRI paradigms. Schizophrenia research, 226, 111–119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohanty, A. , Gitelman, D. R. , Small, D. M. , & Mesulam, M. M. (2008). The spatial attention network interacts with limbic and monoaminergic systems to modulate motivation‐induced attention shifts. Cerebral Cortex, 18(11), 2604–2613. 10.1093/cercor/bhn021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Namboodiri, V. M. K. , & Stuber, G. D. (2020). Interoceptive inception in insula. Neuron, 105(6), 959–960. 10.1016/j.neuron.2020.02.032 [DOI] [PubMed] [Google Scholar]
- Nader, K. , Bechara, A. , & van der Kooy, D. (1997). Neurobiological constraints on behavioral models of motivation. Annual Review of Psychology, 48(1), 85–114. 10.1146/annurev.psych.48.1.85 [DOI] [PubMed] [Google Scholar]
- Naqvi, N. H. , & Bechara, A. (2010). The insula and drug addiction: An interoceptive view of pleasure, urges, and decision‐making. Brain Structure and Function, 214(5‐6), 435–450. 10.1007/s00429-010-0268-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naqvi, N. H. , Gaznick, N. , Tranel, D. , & Bechara, A. (2014). The insula: A critical neural substrate for craving and drug seeking under conflict and risk. Annals of the New York Academy of Sciences, 1316, 53–70. 10.1111/nyas.12415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niv, Y. , Joel, D. , & Dayan, P. (2006). A normative perspective on motivation. Trends in Cognitive Sciences, 10(8), 375–381. 10.1016/j.tics.2006.06.010 [DOI] [PubMed] [Google Scholar]
- Noël, X. , Brevers, D. , & Bechara, A. (2013). A triadic neurocognitive approach to addiction for clinical interventions. Frontiers in Psychiatry, 4, 179. 10.3389/fpsyt.2013.00179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Brien, C. P. , Volkow, N. , & Li, T. K. (2006). What's in a word? Addiction versus dependence in DSM‐V. American Journal of Psychiatry, 163(5), 764–765. 10.1176/ajp.2006.163.5.764 [DOI] [PubMed] [Google Scholar]
- O'Doherty, J. P. (2004). Reward representations and reward‐related learning in the human brain: Insights from neuroimaging. Current Opinion in Neurobiology, 14(6), 769–776. 10.1016/j.conb.2004.10.016 [DOI] [PubMed] [Google Scholar]
- O'Doherty, J. P. , Deichmann, R. , Critchley, H. D. , & Dolan, R. J. (2002). Neural responses during anticipation of a primary taste reward. Neuron, 33(5), 815–826. 10.1016/S0896-6273(02)00603-7 [DOI] [PubMed] [Google Scholar]
- Oldham, S. , Murawski, C. , Fornito, A. , Youssef, G. , Yücel, M. , & Lorenzetti, V. (2018). The anticipation and outcome phases of reward and loss processing: A neuroimaging meta‐analysis of the monetary incentive delay task. Human Brain Mapping, 39(8), 3398–3418. 10.1002/hbm.24184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ostlund, S. B. , & Balleine, B. W. (2007). The contribution of orbitofrontal cortex to action selection. Annals of the New York Academy of Sciences, 1121(1), 174–192. 10.1196/annals.1401.033 [DOI] [PubMed] [Google Scholar]
- Panksepp, J. (2004). Affective neuroscience: The foundations of human and animal emotions. Oxford University Press. [Google Scholar]
- Petzschner, F. H. , Garfinkel, S. N. , Paulus, M. P. , Koch, C. , & Khalsa, S. S. (2021). Computational models of interoception and body regulation. Trends in neurosciences, 44(1), 63–76. 10.1016/j.tins.2020.09.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panksepp, J. , & Biven, L. (2012). The archaeology of mind: Neuroevolutionary origins of human emotions (Norton series on interpersonal neurobiology). WW Norton & Company. [Google Scholar]
- Passingham, R. E. , & Wise, S. P. (2012). The neurobiology of the prefrontal cortex: Anatomy, evolution, and the origin of insight. OUP Oxford. [Google Scholar]
- Robinson, T. E. , & Berridge, K. C. (1993). The neural basis of drug craving: An incentive‐sensitization theory of addiction. Brain Research Reviews, 18(3), 247–291. 10.1016/0165-0173(93)90013-P [DOI] [PubMed] [Google Scholar]
- Robinson, T. E. , Yager, L. M. , Cogan, E. S. , & Saunders, B. T. (2014). On the motivational properties of reward cues: Individual differences. Neuropharmacology, 76, 450–459. 10.1016/j.neuropharm.2013.05.040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salamone, J. D. , & Correa, M. (2002). Motivational views of reinforcement: Implications for understanding the behavioral functions of nucleus accumbens dopamine. Behavioural Brain Research, 137(1–2), 3–25. 10.1016/S0166-4328(02)00282-6 [DOI] [PubMed] [Google Scholar]
- Salamone, J. D. , Correa, M. , Yang, J. H. , Rotolo, R. , & Presby, R. (2018). Dopamine, effort‐based choice, and behavioral economics: Basic and translational research. Frontiers in Behavioral Neuroscience, 12, 52. 10.3389/fnbeh.2018.00052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salamone, J. D. , Cousins, M. S. , & Snyder, B. J. (1997). Behavioral functions of nucleus accumbens dopamine: Empirical and conceptual problems with the anhedonia hypothesis. Neuroscience & Biobehavioral Reviews, 21(3), 341–359. 10.1016/S0149-7634(96)00017-6 [DOI] [PubMed] [Google Scholar]
- Salamone, J. D. , Yohn, S. E. , López‐Cruz, L. , San Miguel, N. , & Correa, M. (2016). Activational and effort‐related aspects of motivation: Neural mechanisms and implications for psychopathology. Brain, 139(5), 1325–1347. 10.1093/brain/aww050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schott, B. H. , Minuzzi, L. , Krebs, R. M. , Elmenhorst, D. , Lang, M. , Winz, O. H. , Seidenbecher, C. I. , Coenen, H. H. , Heinze, H.‐J. , Zilles, K. , Duzel, E. , & Bauer, A. (2008). Mesolimbic functional magnetic resonance imaging activations during reward anticipation correlate with reward‐related ventral striatal dopamine release. Journal of Neuroscience, 28(52), 14311–14319. 10.1523/JNEUROSCI.2058-08.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schultz, W. (1998). Predictive reward signal of dopamine neurons. Journal of Neurophysiology, 80(1), 1–27. 10.1152/jn.1998.80.1.1 [DOI] [PubMed] [Google Scholar]
- Schultz, W. (2015). Neuronal reward and decision signals: From theories to data. Physiological Reviews, 95(3), 853–951. 10.1152/physrev.00023.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schultz, W. , Dayan, P. , & Montague, P. R. (1997). A neural substrate of prediction and reward. Science, 275(5306), 1593–1599. 10.1126/science.275.5306.1593 [DOI] [PubMed] [Google Scholar]
- Schultz, W. , Tremblay, L. , & Hollerman, J. R. (2000). Reward processing in primate orbitofrontal cortex and basal ganglia. Cerebral Cortex, 10(3), 272–283. 10.1093/cercor/10.3.272 [DOI] [PubMed] [Google Scholar]
- Schwabe, L. , & Wolf, O. T. (2010). Socially evaluated cold pressor stress after instrumental learning favors habits over goal‐directed action. Psychoneuroendocrinology, 35(7), 977–986. 10.1016/j.psyneuen.2009.12.010 [DOI] [PubMed] [Google Scholar]
- Seger, C. A. , & Cincotta, C. M. (2005). The roles of the caudate nucleus in human classification learning. Journal of Neuroscience, 25(11), 2941–2951. 10.1523/JNEUROSCI.3401-04.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sescousse, G. , Caldú, X. , Segura, B. , & Dreher, J. C. (2013). Processing of primary and secondary rewards: A quantitative meta‐analysis and review of human functional neuroimaging studies. Neuroscience & Biobehavioral Reviews, 37(4), 681–696. 10.1016/j.neubiorev.2013.02.002 [DOI] [PubMed] [Google Scholar]
- Siep, N. , Roefs, A. , Roebroeck, A. , Havermans, R. , Bonte, M. L. , & Jansen, A. (2009). Hunger is the best spice: An fMRI study of the effects of attention, hunger and calorie content on food reward processing in the amygdala and orbitofrontal cortex. Behavioural Brain Research, 198(1), 149–158. 10.1016/j.bbr.2008.10.035 [DOI] [PubMed] [Google Scholar]
- Silverman, M. H. , Krueger, R. F. , Iacono, W. G. , Malone, S. M. , Hunt, R. H. , & Thomas, K. M. (2014). Quantifying familial influences on brain activation during the monetary incentive delay task: An adolescent monozygotic twin study. Biological Psychology, 103, 7–14. 10.1016/j.biopsycho.2014.07.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon, J. J. , Skunde, M. , Walther, S. , Bendszus, M. , Herzog, W. , & Friederich, H. C. (2016). Neural signature of food reward processing in bulimic‐type eating disorders. Social Cognitive and Affective Neuroscience, 11(9), 1393–1401. 10.1093/scan/nsw049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon, J. J. , Skunde, M. , Wu, M. , Schnell, K. , Herpertz, S. C. , Bendszus, M. , Herzog, W. , & Friederich, H. C. (2015). Neural dissociation of food‐and money‐related reward processing using an abstract incentive delay task. Social cognitive and Affective Neuroscience, 10(8), 1113–1120. 10.1093/scan/nsu162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spear, L. P. (2011). Rewards, aversions and affect in adolescence: Emerging convergences across laboratory animal and human data. Developmental cognitive neuroscience, 1(4), 390–403. 10.1016/j.dcn.2011.08.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stern, P. (2000). Toward a coherent theory of environmentally significant behavior. Journal of Social Issues, 56(3), 407–424. Cambridge university press. [Google Scholar]
- Strack, F. , & Deutsch, R. (2004). Reflection and impulse as determinants of conscious and unconscious motivation. In: Social motivation: Conscious and unconscious processes (pp. 91–112). Cambridge University Press. [Google Scholar]
- Takahashi, Y. K. , Langdon, A. J. , Niv, Y. , & Schoenbaum, G. (2016). Temporal specificity of reward prediction errors signaled by putative dopamine neurons in rat VTA depends on ventral striatum. Neuron, 91(1), 182–193. 10.1016/j.neuron.2016.05.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toates, F. (1994). Comparing motivational systems—An incentive motivation perspective. In Legg, C. R. , & Booth, D. A. (Eds.), Appetite: Neural and behavioural bases (pp. 305–327). Oxford University Press. 10.1093/acprof:oso/9780198547877.003.0013 [DOI] [Google Scholar]
- Tricomi, E. , Balleine, B. W. , & O'Doherty, J. P. (2009). A specific role for posterior dorsolateral striatum in human habit learning. European Journal of Neuroscience, 29(11), 2225–2232. 10.1111/j.1460-9568.2009.06796.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turel, O. , & Bechara, A. (2016). A triadic reflective‐impulsive‐interoceptive awareness model of general and impulsive information system use: Behavioral tests of neuro‐cognitive theory. Frontiers in Psychology, 7, 601. 10.3389/fpsyg.2016.00601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turkeltaub, P. E. , Eden, G. F. , Jones, K. M. , & Zeffiro, T. A. (2002). Meta‐analysis of the functional neuroanatomy of single‐word reading: Method and validation. Neuroimage, 16(3), 765–780. 10.1006/nimg.2002.1131 [DOI] [PubMed] [Google Scholar]
- van der Laan, L. N. , De Ridder, D. T. , Viergever, M. A. , & Smeets, P. A. (2011). The first taste is always with the eyes: A meta‐analysis on the neural correlates of processing visual food cues. Neuroimage, 55(1), 296–303. 10.1016/j.neuroimage.2010.11.055 [DOI] [PubMed] [Google Scholar]
- Volkow, N. D. , Wang, G. J. , Fowler, J. S. , Logan, J. , Gatley, S. J. , Wong, C. , Hitzemann, R. , & Pappas, N. R. (1999). Reinforcing effects of psychostimulants in humans are associated with increases in brain dopamine and occupancy of D2receptors. Journal of Pharmacology and Experimental Therapeutics, 291(1), 409–415. [PubMed] [Google Scholar]
- Warlow, S. M. , & Berridge, K. C. (2021). Incentive motivation: ‘Wanting’ roles of central amygdala circuitry. Behavioural Brain Research, 411, 113376. 10.1016/j.bbr.2021.113376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watson, P. , Wiers, R. W. , Hommel, B. , & De Wit, S. (2014). Working for food you don't desire. Cues interfere with goal‐directed food‐seeking. Appetite, 79, 139–148. 10.1016/j.appet.2014.04.005 [DOI] [PubMed] [Google Scholar]
- Wei, L. , Zhang, S. , Turel, O. , Bechara, A. , & He, Q. (2017). A tripartite neurocognitive model of internet gaming disorder. Frontiers in Psychiatry, 8, 285. 10.3389/fpsyt.2017.00285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson, R. P. , Colizzi, M. , Bossong, M. G. , Allen, P. , Kempton, M. , & Bhattacharyya, S. (2018). The neural substrate of reward anticipation in health: A meta‐analysis of fMRI findings in the monetary incentive delay task. Neuropsychology review, 28(4), 496–506. 10.1007/s11065-018-9385-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo, C. W. , Krishnan, A. , & Wager, T. D. (2014). Cluster‐extent based thresholding in fMRI analyses: Pitfalls and recommendations. Neuroimage, 91, 412–419. 10.1016/j.neuroimage.2013.12.058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeomans, M. R. , & Mobini, S. (2006). Hunger alters the expression of acquired hedonic but not sensory qualities of food‐paired odors in humans. Journal of Experimental Psychology: Animal Behavior Processes, 32(4), 460–466. 10.1037/0097-7403.32.4.460 [DOI] [PubMed] [Google Scholar]
- Young, C. B. , & Nusslock, R. (2016). Positive mood enhances reward‐related neural activity. Social Cognitive and Affective Neuroscience, 11(6), 934–944. 10.1093/scan/nsw012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yousuf, M. , Heldmann, M. , Göttlich, M. , Münte, T. F. , & Doñamayor, N. (2018). Neural processing of food and monetary rewards is modulated by metabolic state. Brain Imaging and Behavior, 12(5), 1379–1392. 10.1007/s11682-017-9811-y [DOI] [PubMed] [Google Scholar]
- Zellner, D. A. , Rozin, P. , Aron, M. , & Kulish, C. (1983). Conditioned enhancement of human's liking for flavor by pairing with sweetness. Learning and Motivation, 14(3), 338–350. 10.1016/0023-9690(83)90021-8 [DOI] [Google Scholar]
- Zhang, J. , Berridge, K. C. , Tindell, A. J. , Smith, K. S. , & Aldridge, J. W. (2009). A neural computational model of incentive salience. PLoS Computational Biology, 5(7), e1000437. 10.1371/journal.pcbi.1000437 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material
Data Availability Statement
All data are available upon request.