

Abstract
Background
An innovative approach has been introduced for identifying and developing novel potent and safe anti-Babesia and anti-Theileria agents for the control of animal piroplasmosis. In the present study, we evaluated the inhibitory effects of Malaria Box (MBox) compounds (n = 8) against the growth of Babesia microti in mice and conducted bioinformatics analysis between the selected hits and the currently used antibabesial drugs, with far-reaching implications for potent combinations.
Methods
A fluorescence assay was used to evaluate the in vivo inhibitory effects of the selected compounds. Bioinformatics analysis was conducted using hierarchical clustering, distance matrix and molecular weight correlation, and PubChem fingerprint. The compounds with in vivo potential efficacy were selected to search for their target in the piroplasm parasites using quantitative PCR (qPCR).
Results
Screening the MBox against the in vivo growth of the B. microti parasite enabled the discovery of potent new antipiroplasm drugs, including MMV396693 and MMV665875. Interestingly, statistically significant (P < 0.05) downregulation of cysteine protease mRNA levels was observed in MMV665875-treated Theileria equi in vitro culture in comparison with untreated cultures. MMV396693/clofazimine and MMV665875/atovaquone (AV) showed maximum structural similarity (MSS) with each other. The distance matrix results indicate promising antibabesial efficacy of combination therapies consisting of either MMV665875 and AV or MMV396693 and imidocarb dipropionate (ID).
Conclusions
Inhibitory and hematology assay results suggest that MMV396693 and MMV665875 are potent antipiroplasm monotherapies. The structural similarity results indicate that MMV665875 and MMV396693 have a similar mode of action as AV and ID, respectively. Our findings demonstrated that MBox compounds provide a promising lead for the development of new antibabesial therapeutic alternatives.
Graphical Abstract

Supplementary Information
The online version contains supplementary material available at 10.1186/s13071-022-05430-4.
Keywords: Babesia microti, Malaria Box, Bioinformatics analysis, MMV396693, MMV665875
Background
Piroplasmosis is a tick-borne parasitic disease causing great economic loss in the livestock industry worldwide [1]. Diminazene aceturate (DA) and imidocarb dipropionate (ID) have been the standard treatments for babesiosis for many years [2]. However, they have major side effects including a long period for tissue removal, toxicity, and in the case of DA, unavailability in certain countries. Furthermore, new research has revealed that some Babesia species such as Babesia gibsoni may develop DA resistance [3, 4]. Consequently, searching for potent new antibabesial drugs in the currently available antiprotozoal libraries may help in filling this empty antibabesial drug pipeline. In this regard, we recently conducted a large-scale screening of Malaria Box (MBox) compounds (n = 400) against the in vitro growth of Babesia bovis, Babesia bigemina, Babesia caballi, and Theileria equi, and 10 potent Medicines for Malaria Venture (MMV) compounds were identified, which showed a wide potential effect against both bovine and equine piroplasm parasites [5, 6]. Since the in vitro effect of these novel candidates does not reveal the effect of host-related factors [7], testing their in vivo inhibitory effect is required. As a result, in this study, the in vivo inhibitory effects of these powerful MMV compounds were investigated against Babesia microti, which infects wild small mammals including rodents as the reservoir host [8, 9], and which has served as a useful experimental model for animal babesiosis research [10, 11]. The compounds with potential in vivo efficacy were then selected to investigate their structural similarities with each other, with the commonly used antibabesial drugs (DA, and ID), and with the recently identified antibabesial drug clofazimine (CF), using bioinformatics analysis. Also, the targets of these MMV compounds in the Babesia parasite were identified using quantitative polymerase chain reaction (qPCR). Thus, the data presented in this study provide the veterinary field novel compounds with potential efficacy against both bovine babesiosis and equine piroplasmosis. Also, this study suggests a combination therapy for possible use in the treatment of human babesiosis.
Methods
In vivo efficacy of MMV compounds and median lethal dose determination
The anti-B. microti effects of eight MMV compounds (MolPort, Latvia) were investigated in this study in 8-week-old female BALB/c mice (CLEA, Japan) using a fluorescence-based SYBR Green I (SGI) (Lonza, USA; 10,000×) assay [10]. Five mice were allocated to each group. All mice were intraperitoneally injected with 1 × 107 B. microti (Munich strain)-infected red blood cells (RBCs), except for one group that remained uninfected to use as a negative control. The level of parasitemia in the infected mice was monitored daily using Giemsa-stained thin blood smears prepared from venous tail blood, and beginning day 4 post-infection (p.i.), when the parasitemia reached approximately 1%, either MMV compounds or control drug (DA, Novartis, Japan) was intraperitoneally administered to the mice for five successive days [10–14]. Double-distilled water, sterile phosphate-buffered saline 1× (PBS 1×), or a sterile PBS 1× and DMSO (dimethylsulfoxide) solution was used for dissolving MMV compounds, with a final concentration of 0.01% to 0.02%, while DA was dissolved in autoclaved physiological saline (0.9% NaCl w/v, pH 7.2) at a dosage rate of 25 mg kg−1. Various doses of the selected MMV compounds were administered (Fig. 1). After the start of treatment, the emitted fluorescence signals were monitored every 2 days until the cessation of parasitemia. Next, the median lethal dose (LD50) value for MMV396693 was determined using a “staircase” method with doses in ascending order ranging from 10 mg kg−1 to 100 mg kg−1 (five mice per dose) following a protocol previously described by Randhawa et al. [15]. The staircase technique is a limit method in which stimuli are presented in ascending and descending order. The direction of the stimulus sequence is reversed when the observer’s response changes. Because it does not deliver stimuli that are far above or below the threshold, this strategy is effective [15].
Fig. 1.

The growth inhibitory effects of MMV compounds with potent in vitro antipiroplasm (bovine Babesia and equine Babesia and Theileria) effects on B. microti in BALB/c mice. a MMV396693. b MMV666093. c MMV665875. d MMV006706. e MMV006787 and MMV007092. f MMV396794 and MMV085203. Each value is the mean and SD of the independent experiments. The number of independent experiments is shown in brackets. P < 0.05 between treated and untreated mice is indicated by asterisks
Hemolytic anemia monitoring
Hematological parameters, including hemoglobin (HGB) levels, RBC counts, and hematocrit (HCT), were monitored every 96 h using a Celltac Alpha MEK-6450 automatic hematology analyzer to assess the efficacy of MMV monotherapy in preventing the progression of anemia in the treated mice as detailed previously [10, 11].
Atom pair fingerprint measurements
DA, ID, CF, and AV atom pair fingerprints (APfp) were utilized to calculate the structural similarity between the MMV compounds with potent anti-B. microti activity (MMV396693 and MMV665875), either with each other or with other antibabesial drugs [16]. The compound identification number (CID) acquired from PubChem for each compound was used to compute the APfp of the two MMV compounds and other antibabesial medicines. The CIDs were entered into ChemMine Web Tools software, which calculated the APfp of all compounds [17]. ChemmineR software was used to analyze the APfp for hierarchical cluster analysis (HCA) [18].
RNA extraction and qPCR
To evaluate the effects of treatment with the potent identified antipiroplasm MMV compound MMV665875 on the messenger RNA (mRNA) level of the target gene, qPCR was used. Babesia bovis (Texas strain) [19, 20] and T. equi (United States Department of Agriculture [USDA] strain) [6, 21] were cultivated in 24-well plates using a microaerophilic, stationary-phase culture system [19, 22] and treated with the IC99 (is approximately 100-fold the IC50 concentration) of the specific potent MMV compound for 8 h. A culture that received no treatment was used as a negative control. RBCs were collected from all cultures and washed with PBS. Then, total RNA was extracted, complementary DNA (cDNA) was synthesized, and the SGI PCR Master Mix (Lonza, USA) was used as detailed in our previous study [23]. Target gene fold alterations in comparison with 18S rRNA [24, 25] were estimated in MMV665875-treated and DMSO-treated B. bovis or T. equi as described previously [26].
The assay was used for amplification of the cysteine protease (CP) (GenBank accession numbers AK441400 for B. bovis [CP2] and XM_004833324.1 for T. equi) gene from RNA extracted from either MMV-treated or DMSO-treated (control) cultures. Specific forward and reverse primers were designed by Primer3Plus software (http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi/) and used to amplify the target genes (Additional file 3: Table S3).
Statistical analysis
The obtained data were analyzed using GraphPad Prism. Differences between the control and treated groups were determined by one-way analysis of variance (ANOVA) and unpaired t-tests [11, 27]. A P-value < 0.05 was considered statistically significant.
Results
Effects of MBox compounds on B. microti growth in mice
The MBox richness as a resource for creating strong, new antipiroplasm agents was discovered through the in vitro screening of 400 MMV compounds from this box against the growth of B. bovis, B. bigemina, T. equi, and B. caballi parasites [5, 6]. An in vivo investigation was conducted utilizing B. microti in a mouse model to further confirm the MMV compounds as antipiroplasm agents. The highest fluorescence values in the positive control group ranged from 2016 to 2922 (Fig. 1). When compared with control mice, 50 mg kg−1 and 10 mg kg−1 MMV396693 significantly reduced fluorescence levels (P < 0.05) (ANOVA: F(1.121, 22.56) = 17.33, P = 0.0002) from days 8 to 16 and from days 8 to 14 p.i., respectively (Fig. 1a). Fluorescence values were significantly lower (ANOVA: F(1.479, 29.58) = 28.09, P < 0.0001) in mice treated with 25 mg kg−1 MMV666093 at days 8, 18, and 20 p.i. as compared with those in the controls (Fig. 1b), while treatment with 10 mg kg−1 MMV665875 exhibited significant inhibition (ANOVA: F(1.101, 22.03) = 19.81, P = 0.0001) of fluorescence signals from day 8 to day 14 p.i. as compared with that in the controls (Fig. 1c). Administration of 30 mg kg−1 MMV006706 led to significant inhibition (ANOVA: F(2.593, 44.09) = 12.99, P < 0.0001) of the emitted fluorescence signals at day 8 p.i. in comparison with B. microti-infected untreated mice (Fig. 1d).
Inhibition percentages in the in vivo growth of B. microti in the area under the curve for all tested drugs are shown in Additional file 3: Table S1. Collectively, 10 mg kg−1 MMV665875 exhibited the highest anti-B. microti efficacy in vivo among the screened MMV compounds, followed by 10 mg kg−1 MMV396693 and 25 mg kg−1 MMV666093 (Additional file 3: Table S1). Treatment with 25 mg kg−1 MMV007092, MMV006787, MMV396794, and MMV085203 resulted in inhibition of 11.64%, 32.01%, 16.59%, and 45.77% at day 12 p.i., respectively (Fig. 1e, f). Treatment with 10 mg kg−1 and 50 mg kg−1 MMV396693 achieved 53.44% and 84.53% inhibition of B. microti growth at day 10 p.i. and 58.78% and 79.36% inhibition at day 12 p.i., respectively. The inhibition rates after treatment with 25 mg kg−1 DA were 78.05% and 79.75% at days 10 and 12 p.i., respectively (Fig. 1a). However, the inhibitory effect of 50 mg kg−1 MMV396693 caused red discoloration of the mice’s urine, with signs of toxicity such as loss of body weight observed in the treated mice at day 8 p.i. Then the mice started to die, and at day 30 p.i., only two mice remained in this treated group. In light of this issue, we ended the experiment for this dose (50 mg kg) −1at day 30 p.i. and performed a pilot study to calculate the LD50 value for MMV396693. The result revealed that the LD50 for MMV396693, when administered intraperitoneally in mice, was 47.59 mg kg−1 (Fig. 2).
Fig. 2.

A plot of log doses versus probits for calculation of the LD50 of MMV396693 administered intraperitoneally. The experiment was repeated twice
Eight days p.i., the rates of B. microti growth inhibition in mice caused by 10 mg kg−1 and 25 mg kg−1 MMV666093 were 28.89% and 36.21%, respectively (Fig. 1b). Treatment with 30 mg kg−1 MMV006706 caused 17.83% inhibition in the growth of B. microti in mice (Fig. 1d). Interestingly, 10 mg kg−1 MMV665875 caused 38.05% and 53.04% inhibition in the growth of B. microti in mice at days 10 and 12 p.i., respectively (Fig. 1c). These results highlight the potential antipiroplasm efficacy of MMV396693 and MMV665875 (both probe-like compounds) when used as monotherapies.
Effect of MBox compounds on anemia
In this study, HGB levels, RBC counts, and HCT values were calculated to track the recovery of mice from hemolytic anemia caused by B. microti after treatment with powerful MMV antipiroplasm drugs. HGB levels and HCT values were restored to normal levels at day 16 p.i. after treatment with 10 mg kg−1 MMV665875 (Fig. 3a, c). Treatment with MMV396693 (10 mg kg−1) demonstrated better results than that with MMV665875, as the former restored normal RBC counts at day 16 p.i., whereas reduced RBC counts were detected in MMV665875-treated mice until day 20 p.i. (Fig. 3b). These data highlight the quick recovery from anemia caused by B. microti infection once treatment with MMV396693 is initiated and confirm the promising antibabesial efficacy of this hit. Unfortunately, intraperitoneal treatment with 50 mg kg−1 MMV396693 caused significant reductions (ANOVA: F(1.577, 9.461) = 11.73, P = 0.0038 for HGB, ANOVA: F(1.808, 10.98) = 12.95, P = 0.0016 for HCT, and ANOVA: F(1.534, 9.207) = 12.05, P = 0.0039 for RBCs) in all hematological parameters from day 8 p.i. until day 28 p.i. in comparison with negative control mice (Additional file 1: Fig. S1).
Fig. 3.
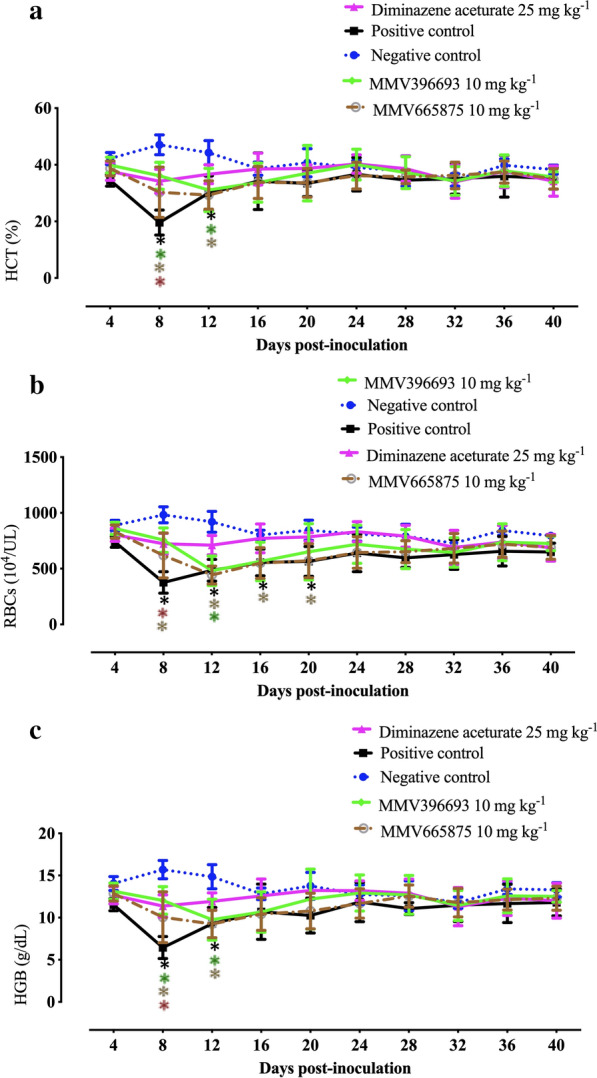
Hematological variables in MBox compound-treated BALB/c mice. a Hematocrit (HCT) levels. b Red blood cell (RBC) counts. d Hemoglobin (HGB) values. Each value is represented as mean and SD of three separate experiments. P < 0.05 between treated and untreated mice is indicated by asterisks
Maximum structural similarity between MBox compounds and antibabesial drugs
MMV396693 (CID: 3618367) exhibited the maximum structural similarity (MSS) to CF (CID: 2794) in the HCA (Fig. 4a). MSS was found between CF (CID: 2794), DA (CID: 5284544), and ID (CID: 9983292) in the same fashion (Fig. 4a). Based on this analysis, AV (CID: 74,989) and MMV665875 (CID: 44522286) both demonstrated MSS with each other (Fig. 4a). Compounds that fall into different clusters have structural differences from one another and from other antibabesial medicines, indicating a possible alternative antibabesial mechanism.
Fig. 4.

Measurements of structural similarity. a Analysis of hierarchical clustering. ChemmineR was used to perform the hierarchical cluster analysis. b A heatmap depicting the distance matrix between diminazene aceturate, imidocarb dipropionate, clofazimine, and atovaquone, which were identified as effective MMV compounds. With Z-score display values, a single-linkage mechanism was used. 3618367 = MMV396693, 44522286 = MMV665875, 5284544 = diminazene aceturate, 9983292 = imidocarb dipropionate, 2794 = clofazimine, and 74989 = atovaquone
Distance matrix between MBox compounds and antibabesial drugs
With both AV and ID, MMV665875 exhibited the lowest distance matrix (LDM) (Fig. 4b and Additional file 3: Table S2). ID showed the LDM with MMV396693 (CID: 3618367) (Fig. 4b and Additional file 3: Table S2). MMV396693 (CID: 3618367) and MMV665875 (CID: 44522286) exhibited very similar molecular weight as that of CF (CID: 2794) (Additional file 2: Fig. S2). Similarity workbench results showed 0.26 AP Tanimoto values between either MMV665875 and AV or MMV665875 and ID, with maximum common substructure (MCS) values of 9 and 10, respectively (Table 1). AP Tanimoto values, which measure the overlap between descriptors for two compounds and are calculated as the ratio between conserved features and the total number of features of each molecule, were relatively high (0.22) between MMV396693 and ID and between MMV665875 and CF (Table 1). Collectively, distance matrix correlation and fingerprint for similarity workbench results highlight the possible potential antibabesial efficacy of either MMV665875 and AV or MMV396693 and ID when administered as a combination therapy.
Table 1.
Similarity workbench between the identified potent MMV compounds and the commonly used and recently identified antibabesial drugs
| MMV396693 & MMV665875 | MMV396693 & AV | MMV396693 & DA | MMV396693 & ID | MMV396693 & CF | MMV665875 & AV | MMV665875 & DA | MMV665875 & ID | MMV665875 & CF | |
|---|---|---|---|---|---|---|---|---|---|
| AP Tanimoto | 0.21 | 0.17 | 0.19 | 0.22 | 0.16 | 0.26 | 0.16 | 0.26 | 0.22 |
| MCS Tanimoto | 0.17 | 0.15 | 0.14 | 0.17 | 0.18 | 0.20 | 0.13 | 0.18 | 0.21 |
| MCS size | 7 | 6 | 7 | 8 | 8 | 9 | 8 | 10 | 11 |
AV atovaquone, DA diminazene aceturate, ID imidocarb dipropionate, CF clofazimine, MCS maximum common substructure
MMV665875 downregulates mRNA of the cysteine protease gene in T. equi
qPCR was utilized to look for probable targets of the newly identified powerful MMV compounds in Babesia parasites. The in vitro treatment of B. bovis with the IC99 of MMV665875 for 8 h downregulated the CP2 gene (Fig. 5a). Of note, no statistically significant difference (ANOVA: F(1, 1) = 11.73, P = 0.4121) was observed in the effects on the CP2 gene in the MMV665875-treated parasites as compared with the control (Fig. 5). Interestingly, a statistically significant difference (ANOVA: F(1, 1) = 322.5, P = 0.0042) was observed in the CP (Fig. 5b) mRNA levels in MMV665875-treated T. equi, with substantial downregulation in comparison with DMSO-treated cultures. The results showed that CP could serve as a target gene of MMV665875 for parasite inhibition in T. equi.
Fig. 5.

mRNA level of the cysteine protease (CP) gene from B. bovis and T. equi cultures treated with MMV665875 at their IC99 values and DMSO (0.1%) for 8 h using qPCR. a B. bovis. b T. equi. P < 0.05 between the fold changes is indicated by asterisks. Three independent experiments were performed
Discussion
The MBox was created to provide a novel avenue for identifying agents to treat apicomplexan parasites [28]. The in vitro inhibitory effects of MBox compounds against 16 protozoa were published in a large, comprehensive dataset [5]. However, the in vivo inhibitory effects, pharmacokinetic/dynamic properties, and LD50 values of most MMV compounds are still unknown. Our recently published in vitro results [5, 6] revealed that 17, 2, and 10 MMV hits were the most interesting with regard to bovine Babesia parasites (B. bovis and B. bigemina), equine Babesia and Theileria parasites (T. equi and B. caballi), and both bovine and equine Babesia and Theileria parasites (B. bovis, B. bigemina, T. equi, and B. caballi), respectively. We recently evaluated the in vivo anti-B. microti effects of the hits with respect to potent anti-bovine Babesia effects (n = 17) [29] and anti-equine piroplasm effects (n = 2) [23]. However, the in vivo inhibitory effects of the hits demonstrating wide potential in vitro effects against both bovine and equine piroplasm parasites (B. bovis, B. bigemina, T. equi and B. caballi) remained to be evaluated. Therefore, in the present study, we evaluated the inhibitory effects of these hits against the growth of B. microti in mice. Compounds with potential in vitro antipiroplasm efficacy were chosen based on three primary criteria: (i) compounds with the highest selectivity indices, (ii) those with the lowest IC50 values, and (iii) those that are easiest to obtain. Out of 10 compounds with potent antipiroplasm activity, MMV396693, MMV666093, MMV006706, and MMV665875 were chosen because they had the lowest IC50 values and the highest selectivity index values. Only MMV085203, MMV396794, MMV006787, and MMV007092 were included in the current in vivo investigation among the remaining MMV compounds (n = 6) due to their availability for commercial purchase.
Out of eight MMV compounds evaluated against the growth of B. microti in mice, MMV396693 and MMV665875 showed potential in vivo activity when used as monotherapy. Treatment of the mice with these two MMV compounds demonstrated that 10 mg kg−1 MMV396693, and 10 mg kg−1 MMV665875 resulted in 81.65% and 77.35% inhibition, respectively, at day 8 p.i. These results are higher than the 21% inhibition for 100 mg kg−1 enoxacin, 15% for 150 mg kg−1 norfloxacin, 23% for 700 mg kg−1 ofloxacin [11], 26% for 125 mg kg−1 pyronaridine tetraphosphate [10], 37% for 50 mg kg−1 thymoquinone [27], 40.38% for 130 mg kg−1 Zingiber officinale rhizome [30], and 65.57% for 200 mg kg−1 myrrh oil [12] monotherapies. On the contrary, the inhibition of B. microti growth in mice from MMV396693 was lower than the 90% inhibition for tafenoquine [31].
Van Voorhis et al. (2016) demonstrated that the CP gene may be the target for MMV665875 to inhibit the growth of Apicomplexa parasites such as Plasmodium falciparum in vitro, while the protozoan target of MMV396693 is still unknown.
Indeed, B. microti and other bovine and equine Babesia/Theileria parasites share biological and parasitological characteristics; for example, all of these parasites are transmitted by ticks, invade RBCs and replicate within them, and cause similar clinical symptoms in dogs, cattle, and equines. Therefore, we evaluated the effects of treatment with the identified antipiroplasm MMV compound on the mRNA level of the target gene in B. bovis and T. equi parasites using qPCR. The IC99 of MMV665875 significantly downregulated the mRNA of the CP gene in the T. equi culture (P < 0.05), indicating that this gene may be the target of this probe-like compound in T. equi. CPs are present in all living organisms and play a critical role in the host cell invasion of many protozoan parasites [32, 33]. For Plasmodium, CPs play a key role in the egression process through their involvement in the degradation of both HGB and erythrocyte cytoskeletal proteins, with the subsequent rupture of the infected erythrocytes [33]. Therefore, CPs represent promising new drug targets for several protozoan diseases, including trypanosomiasis [34], malaria [32], schistosomiasis [35], and leishmaniasis [36]. For piroplasm parasites, a previous in vitro study [37] suggested the presence of CPs in B. bovis. As a result of progress in genomic analysis [38], a subsequent study confirmed the presence of CPs in Babesia and characterized CP in B. bovis. Okubo et al. [37] proposed an essential role of CPs in the invasion of B. bovis to the host RBCs. Ascencio et al. [39] reported that T. equi exhibited extreme expansion of C1A-CP paralogs, which might be functionally associated with the evolution of the schizont stage. However, the exact function of babesial CPs remains uncertain. Although the present study revealed significant downregulation in the mRNA of the CP gene in the T. equi culture treated with MMV665875, more in-depth studies are needed to confirm this possible identified target. Also, further studies are essential to identify the target genes of MMV665875 in B. microti.
Generally speaking, DA and ID are the current standard treatment for animal babesiosis, and over time, antibabesial drug resistance has emerged, especially against DA [3, 4]. Furthermore, the administration of ID is commonly associated with adverse effects, including pain during injection and mild cholinergic signs such as salivation, nasal drip, and brief episodes of vomiting [2]. Simultaneous administration of atropine sulfate is required to reverse the associated cholinergic signs [2]. For human babesiosis, combination therapies are the current regime for treating the infection, with reporting of recovery failure in severe cases and the development of drug-resistant parasites [40]. Therefore, the development of novel combination therapies consisting of low doses of the common animal or human antibabesial drugs with the novel identified MMV compounds might be an effective alternative strategy. Following this pattern, in this study, structural similarities of these MMV compounds either with each other or with the common antibabesial drugs were determined using bioinformatics analysis. In cheminformatics, quantifying the similarity of two molecules is a crucial concept and a common task [41]. Its applications span a variety of domains, the majority of which are connected to medicinal chemistry, such as virtual screening [42]. HCA in the current study revealed MSS between MMV396693 and CF. When the combination was administered to Babesia-infected animal models, the molecular weight correlation heatmap supported the potential promising antibabesial efficacy. Similarly, the distance matrix correlation, similarity workbench fingerprint, and HCA revealed the potential antibabesial efficacy of both MMV665875/AV and MMV396693/ID when administered as combination therapy. Future research is needed to look into the synergistic interactions of these combination therapies against Babesia parasites in vitro and in B. microti-infected mice.
Conclusions
Potent antipiroplasm drugs MMV396693 and MMV665875 were identified in the present study. In vitro treatment of T. equi with the IC99 of MMV665875 for 8 h significantly downregulated the mRNA levels of the CP gene. MMV396693/CF and MMV665875/AV both showed MSS. The distance matrix and similarity workbench fingerprint results highlight the fact that MMV665875 and MMV396693 may have similar modes of action as AV and ID, respectively. This treatment could be used to treat babesiosis in both animals and humans. The targets and mechanisms of action of these drugs may bring novel insights into the biology of Babesia and Theileria.
Supplementary Information
Additional file 1: Figure S1. Anemia monitoring in B. microti-infected mice treated with 50 mg kg−1 MMV396693. a RBC counts. b HGB levels. c Hematocrit values. Each value is the mean and SD of the independent experiments. Asterisks indicate statistically significant (*P < 0.05) difference between treated and untreated mice.
Additional file 2: Figure S2. The molecular weight association between the powerful MMV drugs diminazene aceturate, imidocarb dipropionate, clofazimine, and atovaquone is shown in a heatmap. With Z-scores display values, a single-linkage mechanism was used. 3618367 = MMV396693, 44522286 = MMV665875, 5284544 = diminazene aceturate, 9983292 = imidocarb dipropionate, 2794 = clofazimine, and 74989 = atovaquone.
Additional file 3: Table S1. Inhibitory effects of the tested MMV compounds with potential against the growth of B. microti in mice in comparison with a positive control group. Table S2. Distance matrix correlation between MBox compounds with potential and the currently used antibabesial drugs [diminazene aceturate (DA), imidocarb dipropionate (ID), clofazimine (CF), and atovaquone (AV)]. Table S3. Primers used for determining the mRNA level of the expected target gene from B. bovis and T. equi cultures treated with MMV665875 at their IC99 values and DMSO (0.1%) used for 8 h using a qPCR.
Acknowledgements
The authors would like to thank Dr. Gantuya Sambuu for her technical assistance.
Abbreviations
- MBox
Malaria Box
- B. microti
Babesia microti
- qPCR
quantitative PCR
- MSS
Maximum structural similarity
- AV
Atovaquone
- ID
Imidocarb dipropionate
- DA
Diminazene aceturate
- B. bovis
Babesia bovis
- T. equi
Theileria equi
- MMV
Medicines for Malaria Venture
- CF
Clofazimine
- SGI
SYBR Green I
- DMSO
Dimethylsulfoxide
- HGB
Hemoglobin
- RBC
Red blood cell
- HCT
Hematocrit
- APfp
Atom pair fingerprint
- CID
Compound identification number
- HCA
Hierarchical cluster analysis
- USDA
United States Department of Agriculture
- CP
Cysteine protease
Author contributions
Conceived and designed the experiments: MAR, SAEE, II. Performed the experiments: MAR, SAEE, RE. Analyzed the data: MAR, HBB. Contributed reagents/materials/analysis tools: MAR, HBB, II. Wrote the manuscript: MAR, SAEE, II. All authors reviewed the manuscript.
Funding
This study is supported financially by the Medicines for Malaria Venture Malaria Box Challenge Grant. Mohamed Abdo Rizk is supported by a research grant fellowship for young scientists from the Japan Society for the Promotion of Science (JSPS) (ID no. P18091).
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declarations
Ethics approval and consent to participate
The Animal Care and Use Committee and the Biological Safety Committee of Obihiro University of Agriculture and Veterinary Medicine in Hokkaido, Japan, authorized all of the experiments in this work (Approval no. 27-65, and pathogen experiment ID for Babesia microti: 201709-05).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Mohamed Abdo Rizk and Shimaa Abd El-Salam El-Sayed contributed equally to this work
Contributor Information
Mohamed Abdo Rizk, Email: dr_moh_abdo2008@mans.edu.eg.
Hanadi B. Baghdadi, Email: hbbaghdadi@iau.edu.sa
Shimaa Abd El-Salam El-Sayed, Email: igarcpmi@obihiro.ac.jp, Email: Shimaa_a@mand.edu.eg.
Rasha Eltaysh, Email: rasha_mans@yahoo.com.
References
- 1.Suarez CE, Alzan HF, Silva MG, Rathinasamy V, Poole WA, Cooke BM. Unravelling the cellular and molecular pathogenesis of bovine babesiosis: is the sky the limit. Int J Parasitol. 2019;49:183–97. doi: 10.1016/j.ijpara.2018.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mosqueda J, Olvera-Ramirez A, Aguilar Tipacamu G, Canto GJ. Current advances in detection and treatment of babesiosis. Curr Med Chem. 2012;19:1504–18. doi: 10.2174/092986712799828355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hwang SJ, Yamasaki M, Nakamura K, Sasaki N, Murakami M, Wickramasekara Rajapakshage BK, et al. Development and characterization of a strain of Babesia gibsoni resistant to diminazene aceturate in vitro. J Vet Med Sci. 2010;72:765–71. doi: 10.1292/jvms.09-0535. [DOI] [PubMed] [Google Scholar]
- 4.Yamasaki M, Watanabe N, Idaka N, Yamamori T, Otsuguro K, Uchida N, et al. Intracellular diminazene aceturate content and adenosine incorporation in diminazene aceturate-resistant Babesia gibsoni isolate in vitro. Exp Parasitol. 2017;183:92–8. doi: 10.1016/j.exppara.2017.10.016. [DOI] [PubMed] [Google Scholar]
- 5.Van Voorhis WC, Adams JH, Adelfio R, Ahyong V, Akabas MH, Alano P, et al. Open source drug discovery with the malaria box compound collection for neglected diseases and beyond. PLoS Pathog. 2016;12:e1005763. doi: 10.1371/journal.ppat.1005763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rizk MA, El-Sayed SAE, El-Khodery S, Yokoyama N, Igarashi I. Discovering the in vitro potent inhibitors against Babesia and Theileria parasites by repurposing the Malaria Box: a review. Vet Parasitol. 2019;274:108895. doi: 10.1016/j.vetpar.2019.07.003. [DOI] [PubMed] [Google Scholar]
- 7.Bennett TN, Paguio M, Gligorijevic B, Seudieu C, Kosar AD, Davidson E, et al. Novel, rapid, and inexpensive cell-based quantification of antimalarial drug efficacy. Antimicrob Agents Chemother. 2004;48:1807–10. doi: 10.1128/AAC.48.5.1807-1810.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vannier E, Gewurz BE, Krause PJ. Human babesiosis. Infect Dis Clin North Am. 2008;22:469–88 viii–ix. doi: 10.1016/j.idc.2008.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tołkacz K, Bednarska M, Alsarraf M, Dwużnik D, Grzybek M, Welc-Falęciak R, et al. Prevalence, genetic identity and vertical transmission of Babesia microti in three naturally infected species of vole, Microtus spp. (Cricetidae) Parasit Vector. 2017;10:66. doi: 10.1186/s13071-017-2007-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rizk MA, El Sayed SAE, AbouLaila M, Eltaysh R, Yokoyama N, Igarashi I. Performance and consistency of a fluorescence-based high-throughput screening assay for use in Babesia drug screening in mice. Sci Rep. 2017;7:2774. doi: 10.1038/s41598-017-13052-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rizk MA, AbouLaila M, El Sayed SAE, Guswanto A, Yokoyama N, Igarashi I. Inhibitory effects of fluoroquinolone antibiotics on Babesia divergens and Babesia microti, blood parasites of veterinary and zoonotic importance. Infect Drug Resist. 2018;11:1605–15. doi: 10.2147/IDR.S159519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.AbouLaila M, El Sayed SAE, Omar MA, Al-Aboody MS, Aziz ARA, Abdel Daim MM, et al. Myrrh oil in vitro inhibitory growth on bovine and equine piroplasm parasites and Babesia microti of Mice. Pathogens. 2020 doi: 10.3390/pathogens9030173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Aboulaila M, Munkhjargal T, Sivakumar T, Ueno A, Nakano Y, Yokoyama M, et al. Apicoplast-targeting antibacterials inhibit the growth of Babesia parasites. Antimicrob Agents Chemother. 2012;56:3196–206. doi: 10.1128/AAC.05488-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Salama AA, AbouLaila M, Terkawi MA, Mousa A, El-Sify A, Allaam M, et al. Inhibitory effect of allicin on the growth of Babesia and Theileria equi parasites. Parasitol Res. 2014;113:275–83. doi: 10.1007/s00436-013-3654-2. [DOI] [PubMed] [Google Scholar]
- 15.Randhawa MA. Calculation of LD50 values from the method of Miller and Tainter, 1944. J Ayub Med Coll Abbottabad. 2009;21:184–5. [PubMed] [Google Scholar]
- 16.O’Boyle NM, Sayle RA. Comparing structural fingerprints using a literature based similarity benchmark. J Cheminformatics. 2016;8:2206–19. doi: 10.1186/s13321-016-0148-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Backman TW, Cao Y, Girke T. ChemMine tools: an online service for analyzing and clustering small molecules. Nucleic Acids Res. 2011;39:W486–91. doi: 10.1093/nar/gkr320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cao Y, Charisi A, Cheng LC, Jiang T, Girke T. ChemmineR: a compound mining framework for R. Bioinformatics. 2008;24:1733–44. doi: 10.1093/bioinformatics/btn307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Rizk MA, El Sayed SA, Terkawi MA, Youssef MA, El Said el Sel S, Elsayed G, et al. Optimization of a fluorescence-based assay for large-scale drug screening against Babesia and Theileria parasites. PLoS ONE. 2015;10:5276. doi: 10.1371/journal.pone.0125276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rizk MA, El-Sayed SAE-S, Igarashi I. Effects of methanolic extract from turmeric (Curcuma longa) against the in vitro multiplication of several Babesia species and Theileria equi. Parasitologia. 2021;1:188–96. 10.3390/parasitologia1040020. [DOI]
- 21.Omar MA, Salama A, Elsify A, Rizk MA, Al-Aboody MS, AbouLaila M, et al. Evaluation of in vitro inhibitory effect of enoxacin on Babesia and Theileria parasites. Exp Parasitol. 2016;161:62–67. doi: 10.1016/j.exppara.2015.12.016. [DOI] [PubMed] [Google Scholar]
- 22.Rizk MA, El Sayed SAE, AbouLaila M, Yokoyama N, Igarashi I. Evaluation of the inhibitory effect of N-acetyl-L-cysteine on Babesia and Theileria parasites. Exp Parasitol. 2017;179:43–8. doi: 10.1016/j.exppara.2017.06.003. [DOI] [PubMed] [Google Scholar]
- 23.Rizk MA, El-Sayed SAE, Eltaysh R, Igarashi I. MMV020275 and MMV020490, promising compounds from malaria box for the treatment of equine piroplasmosis. Ticks Tick Borne Dis. 2022;13:101904. doi: 10.1016/j.ttbdis.2022.101904. [DOI] [PubMed] [Google Scholar]
- 24.Tuvshintulga B, AbouLaila M, Sivakumar T, Tayebwa DS, Gantuya S, Naranbaatar K, et al. Chemotherapeutic efficacies of a clofazimine and diminazene aceturate combination against piroplasm parasites and their AT-rich DNA-binding activity on Babesia bovis. Sci Rep. 2017;7:13888. doi: 10.1038/s41598-017-14304-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kim CM, Blanco LB, Alhassan A, Iseki H, Yokoyama N, Xuan X, et al. Diagnostic real-time PCR assay for the quantitative detection of Theileria equi from equine blood samples. Vet Parasitol. 2008;151:158–63. doi: 10.1016/j.vetpar.2007.10.023. [DOI] [PubMed] [Google Scholar]
- 26.Livak KJSTD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods. 2001;25:402–8. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 27.El Sayed SAE, Rizk MA, Yokoyama N, Igarashi I. Evaluation of the in vitro and in vivo inhibitory effect of thymoquinone on piroplasm parasites. Parasit Vector. 2019;12:37. doi: 10.1186/s13071-019-3296-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Spangenberg T, Burrows JN, Kowalczyk P, McDonald S, Wells TN, Willis P. The open access malaria box: a drug discovery catalyst for neglected diseases. PLoS ONE. 2013;8:2906. doi: 10.1371/journal.pone.0062906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Rizk MA, El-Sayed SAE, Eltaysh R, Igarashi I. In vivo antibabesial activity and bioinformatic analysis of compounds derived from the medicines for malaria venture box against Babesia microti. Mol Biochem Parasitol. 2022;247:111444. doi: 10.1016/j.molbiopara.2021.111444. [DOI] [PubMed] [Google Scholar]
- 30.Rizk MA, El-Sayed SAE, Igarashi I. Evaluation of the inhibitory effect of Zingiber officinale rhizome on Babesia and Theileria parasites. Parasitol Int. 2021;85:102431. doi: 10.1016/j.parint.2021.102431. [DOI] [PubMed] [Google Scholar]
- 31.Mordue DG, Wormser GP. Could the drug tafenoquine revolutionize treatment of Babesia microti Infection? J Infect Dis. 2019;220:442–447. doi: 10.1093/infdis/jiz119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Verma SDR, Pandey KC. Cysteine proteases: modes of activation and future prospects as pharmacological targets. Front Pharmacol. 2016;7:107. doi: 10.3389/fphar.2016.00107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kappmeyer LS, Thiagarajan M, Herndon DR, Ramsay JD, Caler E, Djikeng A, et al. Comparative genomic analysis and phylogenetic position of Theileria equi. BMC Genom. 2012;13:603. doi: 10.1186/1471-2164-13-603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Engel JC, Doyle PS, Hsieh I, McKerrow JH. Cysteine protease inhibitors cure an experimental Trypanosoma cruzi infection. J Exp Med. 1998;188:725–34. doi: 10.1084/jem.188.4.725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wasilewski MM, Lim KC, Phillips J, McKerrow JH. Cysteine protease inhibitors block schistosome hemoglobin degradation in vitro and decrease worm burden and egg production in vivo. Mol Biochem Parasitol. 1996;81:179–89. doi: 10.1016/0166-6851(96)02703-x. [DOI] [PubMed] [Google Scholar]
- 36.Das L, Datta N, Bandyopadhyay S, Das PK. Successful therapy of lethal murine visceral leishmaniasis with cystatin involves up-regulation of nitric oxide and a favorable T cell response. J Immunol. 2001;166:4020–8. doi: 10.4049/jimmunol.166.6.4020. [DOI] [PubMed] [Google Scholar]
- 37.Okubo K, Yokoyama N, Govind Y, Alhassan A, Igarashi I. Babesia bovis: effects of cysteine protease inhibitors on in vitro growth. Exp Parasitol. 2007;117:214–7. doi: 10.1016/j.exppara.2007.04.009. [DOI] [PubMed] [Google Scholar]
- 38.Yamagishi J, Wakaguri H, Yokoyama N, Yamashita R, Suzuki Y, Xuan X, et al. The Babesia bovis gene and promoter model: an update from full-length EST analysis. BMC Genom. 2014;15:678. doi: 10.1186/1471-2164-15-678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ascencio ME, Florin Christensen M, Mamoun CB, Weir W, Shiels B, Schnittger L. Cysteine proteinase C1A paralog profiles correspond with phylogenetic lineages of pathogenic piroplasmids. Vet Sci. 2018 doi: 10.3390/vetsci5020041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zintl AMG, Skerrett HE, Taylor SM, Gray JS. Babesia divergens, a bovine blood parasite of veterinary and zoonotic importance. Clin Microbiol Rev. 2003;16:622–636. doi: 10.1128/CMR.16.4.622-636.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bender A, Glen RC. Molecular similarity: a key technique in molecular informatics. Org Biomol Chem. 2004;2:3204–18. doi: 10.1039/B409813G. [DOI] [PubMed] [Google Scholar]
- 42.Eckert H, Bajorath J. Molecular similarity analysis in virtual screening: foundations, limitations and novel approaches. Drug Discov Today. 2007;12:225–33. doi: 10.1016/j.drudis.2007.01.011. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Figure S1. Anemia monitoring in B. microti-infected mice treated with 50 mg kg−1 MMV396693. a RBC counts. b HGB levels. c Hematocrit values. Each value is the mean and SD of the independent experiments. Asterisks indicate statistically significant (*P < 0.05) difference between treated and untreated mice.
Additional file 2: Figure S2. The molecular weight association between the powerful MMV drugs diminazene aceturate, imidocarb dipropionate, clofazimine, and atovaquone is shown in a heatmap. With Z-scores display values, a single-linkage mechanism was used. 3618367 = MMV396693, 44522286 = MMV665875, 5284544 = diminazene aceturate, 9983292 = imidocarb dipropionate, 2794 = clofazimine, and 74989 = atovaquone.
Additional file 3: Table S1. Inhibitory effects of the tested MMV compounds with potential against the growth of B. microti in mice in comparison with a positive control group. Table S2. Distance matrix correlation between MBox compounds with potential and the currently used antibabesial drugs [diminazene aceturate (DA), imidocarb dipropionate (ID), clofazimine (CF), and atovaquone (AV)]. Table S3. Primers used for determining the mRNA level of the expected target gene from B. bovis and T. equi cultures treated with MMV665875 at their IC99 values and DMSO (0.1%) used for 8 h using a qPCR.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.