

Abstract
Introduction
Hyaluronic acid (HA) plays an important role in cellular and extracellular matrix (ECM) homeostasis. Recent studies demonstrate that low molecular weight (MW) HA has pro‐inflammatory characteristics while high MW HA is considered anti‐inflammatory and regenerative. In formulating a topical HA product, the possibility of creating a focused high MW HA technology was posed, combining external surface high MW HA constituents with active agents promoting fibroblast production of high MW in the depths of the dermis.
Methods
Human dermal fibroblasts and keratinocytes were treated with various agents, and RNA sequencing (RNA‐seq) was conducted to identify genes involved in HA synthesis. HA production by fibroblasts was assessed by collecting the culture supernatant, concentrating the protein, and conducting polyacrylamide gel electrophoresis (PAGE). The gel was stained with Stains‐All to identify bands relative to known HA products of different MWs. Subsequently, the supernatants were treated with hyaluronidase to confirm the bands corresponded to HA.
Results
The RNA‐seq results revealed a variety of agents upregulated HA‐related genes. However, a potent upregulation of HA synthesis gene was observed by hexapeptide‐11 in the keratinocytes and a newly identified proprietary octapeptide in the fibroblasts. PAGE demonstrated not only robust production of HA by octapeptide, but significantly, the HA produced was ~2 Mega Daltons in size. Octapeptide was the most potent stimulator among the tested agents.
Conclusion
Comprehensive in vitro testing identified a group of active agents that stimulated high MW HA production. This novel approach to HA topical application with exclusively high MW HA production should maximize hydration capacity while encouraging regenerative activity within the ECM. Multi‐center trials are underway.
Keywords: high molecular weight HA, hyaluronic acid, hydration, octapeptide
1. INTRODUCTION
Hyaluronic acid (HA) plays an important role in cell turnover, metabolism, movement and differentiation, tissue repair, hydration, nutrient exchange, and protection against free radical damage. 1 Its maximum size reaches 107 Da. At the body's pH, it is one of the most highly charged molecules in biology, which provides HA with some of its unique qualities. A large body of water surrounds the molecule attempting to neutralize that negative charge and increasing the original volume by 1000–10 000 times. 1 It is this quality that provides the hydrating function of HA, promoting the expansion of tissues thus opening spaces for cell movement. Because water molecules surround HA, the charges repel each other, thereby causing its “slippery” viscosity and ability to hold shape. 1
Considering the biology of HA, it is dynamic and rapid. A 70 kg individual has approximately 15 g of HA, 50% of which is skin associated. Remarkably, a third of total HA turns over daily. HA is reported to decrease with aging, but HA content, in fact, remains constant with age. The difference is that the HA becomes increasingly tissue bound and more resistant to extraction. This HA encased within tissue proteins may be limited in its hydrating capacity. 1 In the skin, HA is located and produced in both the dermis and epidermis, with the greater proportion residing in the dermis. Epidermal HA is more loosely associated and is more easily extracted from tissue. Skin HA has a very rapid turnover, with a half‐life of 1–2 days in the epidermis. The turnover rate in the dermis is similar, with catabolism occurring in the liver and lymph nodes, following lymphatic drainage. 1 In the basal layer, HA is predominantly intracellular and involved in mitotic events, while extracellular HA in the upper layers of the epidermis is involved in barrier disassociation and the sloughing of cells. The HA‐bound water in both the dermis and the epidermis is critical for skin hydration. 1 The most predominant receptor for HA is CD44. 2
Many topical preparations use combinations of high and low molecular weight (MW) HA to promote skin hydration. However, in the wound‐healing literature low MW HA is known to be pro‐inflammatory and could be problematic when treating the skin. 3 , 4 Low molecular weight HA promotes the production of inflammatory mediators, including nitric oxide synthase 2 (nos2), tumor necrosis factor (TNF), interleukin 12 (IL12b), and cluster of differentiation‐80 (CD80). 4 High MW HA inhibits the production of pro‐inflammatory mediators. 4 High MW HA (>106 Da) is considered a sign of normal healthy tissues, while fragmented HA is a reflection of tissues under stress. 1 There is an intrinsic need to ensure that all HA formulatory materials do not contain short HA chains that could stimulate an inflammatory response, thus limiting regenerative functions. 1 However, even though high MW HA is very efficient in treating the skin surface, it cannot penetrate the skin depth to the dermis. Therefore, in designing a topical HA product, we sought to combine a high MW HA to work on the surface together with agents that stimulate dermal fibroblasts to produce higher MW HA in the depths of the skin. We have identified such actives that do just that.
Optimization of delivery and biological effect was achieved by combining the studied compounds described below. Incorporating these actives with a liposome system ensure effective delivery of select actives.
2. MATERIALS AND METHODS
2.1. Cell culture
Human adult dermal fibroblasts and keratinocytes were purchased from ZenBio (Durham, NC) and cultured in fibroblast and keratinocyte media, respectively (ZenBio). The cells were plated in 6‐well or 48‐well plates (Greiner) depending on the assay. The cells were maintained at 37°C in a 5% CO2 incubator.
2.2. Cell treatments for RNA‐seq
The fibroblasts and keratinocytes were seeded into 48‐well plates and left to adhere for 48 h. Then, the cells were exposed to the following compounds for 24 h: Lactoferrin (500 μg/ml), TCVRRAF (100 μg/ml) (amino acid sequence extracted from Lactoferrin), Tri‐peptide‐1 –100 ppm (2.9 μg/ml), Hexapeptide 12, 100 ppm (2.9 μg/ml), TriHex combination 200 ppm (2.9 μg/ml each), Hexapeptide 11 (100 μg/ml), Tranexamic acid 5% (500 μg/ml), Octapeptide (100 μg/ml)—proprietary peptide designed by R&D at Alastin (Alastin …a Galderma Company), Phosphatidylserine 500 μg/ml, or Cannabidiol (CBD) 100 μg/ml. The control cells were left untreated. The compounds were resuspended in the appropriate vehicle at pre‐determined concentrations.
2.3. RNA lysate preparation
After 24 h of compound exposure, the media was removed, the cells were washed 1× with phosphate‐buffered saline (PBS). A total of 100ul of RNA lysis buffer (Takara Bio Cat Num 635013, “10× RNA lysis buffer,” diluted to 1×) were added to the well and mixed thoroughly by trituration. The slurry was added to an RNAse‐free microcentrifuge tube and immediately frozen at −30°C.
2.4. RNA‐seq
All the RNA samples were shipped frozen on dry ice to MedGenome (Foster City, CA) for RNA extraction, library construction and sequencing to 25 M paired end 100bp reads per sample. Differentially expressed genes were identified, and pathway enrichment was assessed using the reactome pathway. 5
2.5. Selection of agents for HA production and size assessment in vitro
Using agents identified by RNA‐seq as novel HA production stimulants in the fibroblasts, together with other active agents previously determined to stimulate HA production, the production of HA by fibroblasts was confirmed and the MW/size of the produced HA was determined. The components selected are shown in Table 1. The rationale for the selection of these agents is described here.
Octapeptide (proprietary peptide)—upregulates Hyaluronic Acid Synthase‐2 (HAS2) gene expression in fibroblasts (based on our RNA‐seq data)
Lactoferrin—has wound‐healing attributes, promotes fibroblast proliferation, and increases HA secretion 6 , 7
Syn‐Hycan—synthetic tripeptide that stimulates HA. Published evidence suggests that a synthetic tripeptide tetradecyl aminobutyroylvalylaminobutyric urea trifluoroacetate restores facial skin volume by stimulating HA synthesis. 8 Syn‐Hycan increases HA and CD44 in the skin in vitro and ex vivo. 8
Phosphatidylserine—promotes HA synthesis. 9
Sodium Hyaluronate Crosspolymer (Hyalasome)—extremely high MW synthetic HA with exceptionally high water‐binding capacity resulting in excellent moisturizing abilities. 10
Tremella fuciformis extract derived from an edible mushroom—provides high levels of moisture from as a natural HA stimulant and has antioxidant properties 11 , 12
Hydroxymethoxyphenyl decanone—HA booster, antioxidant, and anti‐irritant. It has been demonstrated to stimulate the dermal and epidermal hyaluronic acid levels by 259% and 198% versus placebo, respectively, in an ex vivo human skin model. 13
Hexapeptide‐11—upregulates HAS2 and potently downregulates HA reducing enzyme hyaluronidase 2 (HYAL2) in keratinocytes (based on our RNA‐seq data)
TABLE 1.
Compounds used to treat fibroblasts for HA production assessment by PAGE
| Compound | Concentration | |
|---|---|---|
| 1 | Octapeptide | 100 µg/ml |
| 2 | Lactoferrin | 500 µg/ml |
| 3 | SynHycan | 500 µg/ml |
| 4 | Phosphatidylserine | 500 µg/ml |
| 5 | Hyalasome | 500 µg/ml |
| 6 | Tremella | 500 µg/ml |
| 7 | Hydroxymethoxphenly decanone | 250 µg/ml |
| 8 | Aquaxyl | 500 µg/ml |
| 9 | Mix of all compounds | 12.5% of the above concentrations |
| 10 | Mix of all compounds | 40% of the above concentrations |
| 11 | No treatment |
2.6. HA production and size assessment in vitro
Fibroblasts were cultured as described above until near confluence in 6‐well plates. Then, the cells were treated with the compounds shown in Table 1. After 72 h, 100μl of media was collected from each treatment condition. The media was concentrated in a SpeedVac concentrator to a final volume of ~10μl. All 10 μl from each condition was loaded onto an acrylamide gel (NuPAGE 4%–12% Bis‐Tris Protein Gel; Invitrogen). Sodium Hyaluronate, Research Grade, (HA2 M) from Lifecore Biomedical was reconstituted as recommended by the manufacturer and served as a MW reference, representing high MW HA. The gel was run at 200 V for 3 h. Gel buffer was exchanged with fresh room temperature buffer every 20 min to avoid overheating. The gel was stained using Stains‐All (Millipore Sigma) and destained according to the manufacturer's protocol. A band running at the same size of the HA2 M band indicated the production of high MW HA.
2.6.1. Confirmation of HA
Fibroblasts were cultured as described above until near confluence in 6‐well plates. Then, the cells were treated with the compounds shown in Table 1. After 72 h, 100μl of media was collected from each treatment condition. The media was then subjected to hyaluronidase enzyme treatment (1 mg/ml) at 37°C for 2 h. One condition was left non‐digested (octapeptide at 100 μg/ml). The samples were run on the acrylamide gel, stained, and destained as described above. The absence of the band after digestion indicated that the band identified in the prior step was HA.
3. RESULTS
3.1. RNA‐seq reveals 2 peptides that regulate HA gene expression
Examination of the differentially expressed genes from the fibroblast RNA‐seq analysis revealed that HAS2 was significantly upregulated by Octapeptide (Octa) (Figure 1). For the keratinocytes, hexapeptide 11 (Hex11) also upregulated HAS2 and showed a potent downregulation of HYAL2 (Figure 2A,B). The early growth response 3 (EGR3) gene was also upregulated in keratinocytes treated with Hex11 (Figure 3). EGR3 is a transcription factor specifically present in the granular layer of the epidermis and is the gene that is responsible for the formation of the skin barrier. 14 Selectively increasing EGR3 expression in keratinocytes strengthens the skin barrier. 14
FIGURE 1.
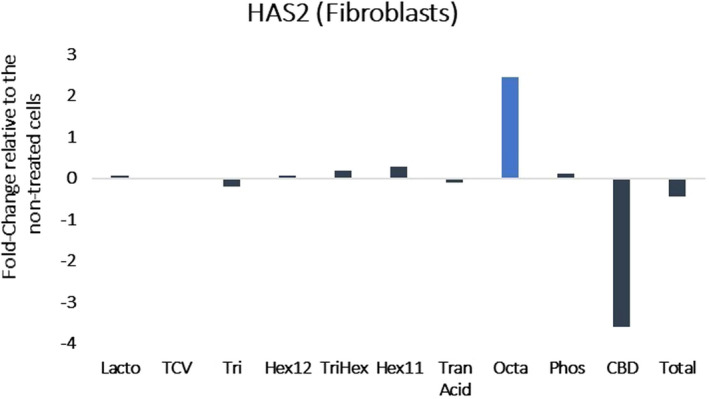
Results of RNA‐seq presented as the fold change in gene expression relative to the non‐treated cells. (HAS2: hyaluronic acid synthase 2). Octapeptide upregulates hyaluronic acid synthase gene expression in dermal fibroblasts
FIGURE 2.
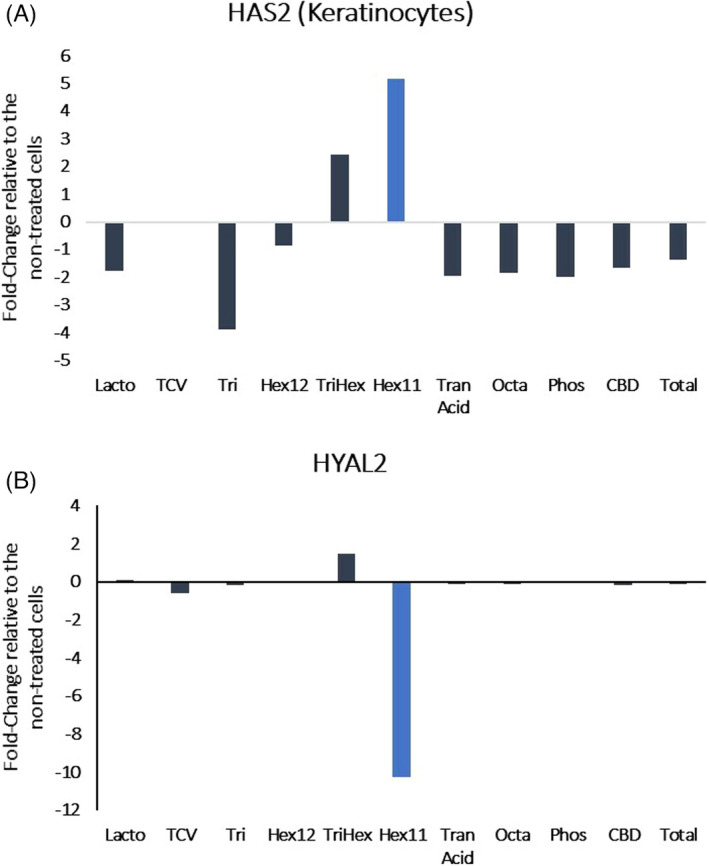
Results of RNA‐seq presented as the fold change in gene expression relative to the non‐treated cells (HAS2: hyaluronic acid synthase 2 and HYAL2: hyaluronidase 2). Hexapeptide‐11 regulates hyaluronic acid‐related gene expression in keratinocytes
FIGURE 3.
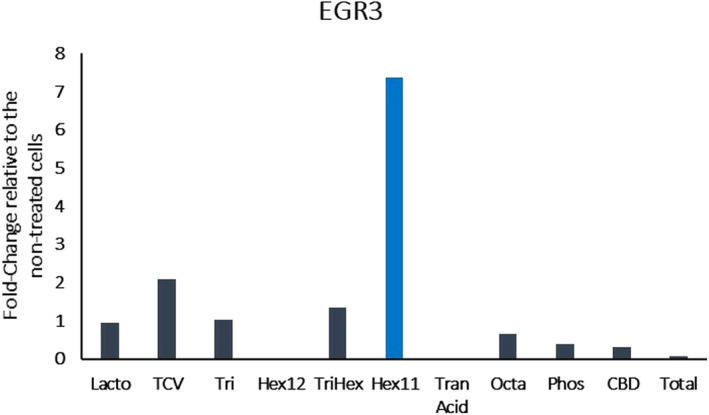
Results of RNA‐seq presented as the fold change in gene expression relative to the non‐treated cells (EGR3: early growth response 3). Hexapeptide‐11 upregulates early growth response 3 gene expression in keratinocytes
3.2. Assessing fibroblast HA production and size in vitro
Determining whether a compound stimulates fibroblasts to secrete HA is important. Thus, based on our RNA‐seq data, we treated fibroblasts with Octa and other compounds, including Lactoferrin, SynHycan, Phosphatidylserine, Hyalasome, Tremella, Symdecanox, Aquaxyl, or a combination of all of these, as described in the methods. Then, we assessed HA production. All the treatments stimulated high MW HA (~2 MegaDaltons (MDa)) secretion (Figure 4).
FIGURE 4.

Gel diffusion study results—the lane numbers are indicated by the treatments described above, and lane 12 is the MW reference, indicating that octapeptide stimulates HA production in dermal fibroblasts at 2 MegaDaltons (2 MDa)
Furthermore, the presence of HA was validated by digesting the fibroblast‐derived supernatants with hyaluronidase and observing the disappearance of the bands (Figure 5).
FIGURE 5.

HA production confirmation by hyaluronidase treatment. For these eight conditions, the supernatants were subjected to hyaluronidase. In addition, one group of cells treated with Octapeptide was not treated with hyaluronidase. Next, the supernatants were concentrated, run on an acrylamide gel, stained and imaged. The lane numbers are indicated by the treatments described above, and lane 10 is the MW reference, indicating HA at 2 MDa
Importantly, Octapeptide alone induced the strongest HA band. Thus, we examined it at 2 doses and revealed a dose‐dependent increase (Figure 6). Thus, these compounds are major stimulators of high MW HA in fibroblasts, with the likelihood that Octapeptide is the strongest driver of this production.
FIGURE 6.
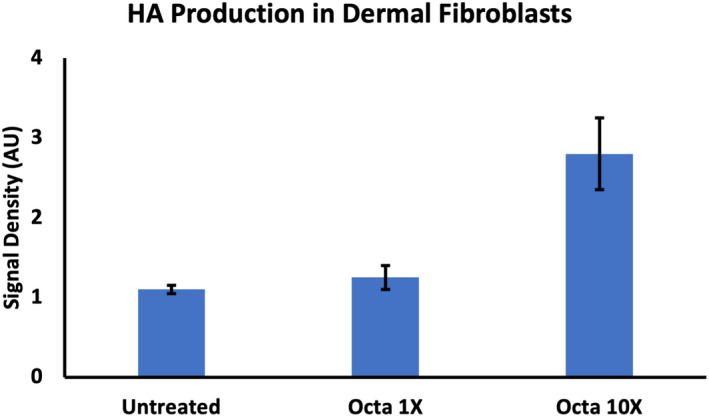
Human dermal fibroblasts were treated with Octapeptide (1× and 10×) or left untreated and then run on an acrylamide gel, stained and imaged. The experiment was conducted 4 times, and the density of the bands from each supernatant was quantified (arbitrary units (AU)) using ImageJ. The data represent the mean ± standard deviation of the band densities. Octapeptide stimulates High MW HA in a dose–response manner in dermal fibroblasts
4. DISCUSSION
HA is an essential ECM constituent providing a milieu that optimizes cellular function and turnover and contributes to overall matrix homeostasis. The hydrating function of HA promotes cellular health on the surface of the skin as well as in the depths of the dermis by providing the ECM with nutrients and allowing the free flow of cells. 1 Recent studies have focused on the functional aspects of HA that relate to its MW.
Many properties of HA depend on its MW—the higher the MW, the stronger the water containing properties and the stronger the resistance to the degradation of hyaluronidase. 15 High molecular weight HA displays anti‐inflammatory and immunosuppressive properties, whereas low molecular weight HA is a potent pro‐inflammatory molecule. 3 In addition, further studies have demonstrated that macrophages undergo phenotypic changes dependent on the MW of hyaluronan that correspond to either (1) a pro‐inflammatory response to low MW HA or (2) a pro‐resolving response to high MW HA. 4 Most of these anti‐inflammatory properties are attributable to interactions of high MW HA and CD44, the major cell‐surface HA‐binding protein. 16
Based on these observations and studies, it is apparent that under normal circumstances in unwounded skin, proper hydration is required for skin health maintenance. Thus, the ability to apply high MW HA on the surface of the skin and at the same time stimulate keratinocyte and fibroblast high MW HA production within the depths of the skin constitutes an optimized skin hydration regimen. Indeed, our study demonstrated that certain compounds, including one novel factor (Octapeptide), have the capacity to stimulate high MW HA production.
Initially, using the power of RNA‐seq technology, our gene expression studies enabled us to identify hexapeptide‐11 as potential potent stimulator of HA in keratinocytes. In addition, an added benefit to skin barrier strength and integrity was the demonstration of the upregulation of EGR3 gene expression by hexapeptide‐11 in the keratinocytes. Indeed, hexapeptide‐11 is a proven powerful regulator in the skin as it upregulates autophagy, encouraging lipid droplet breakdown. Moreover, in vitro studies reveal that hexapeptide‐11 upregulates macrophage recruitment to damaged fat cells, and in vivo trials confirm that it increases and hastens fat volume reduction. 17 , 18 , 19 While the activation of keratinocytes by hexapeptide‐11 is exciting, the fibroblasts are a major source of continued HA production. The newly identified octapeptide (now named octapeptide‐45), as a proprietary peptide, showed potent upregulation of an HA synthesis gene by dermal fibroblasts, and this is an exciting development.
To investigate whether the gene expression results led to functional changes in the fibroblasts under the influence of octapeptide, we assessed HA production and size using a PAGE system. Octapeptide and other agents previously studied as HA stimulators not only stimulated the production of HA, but the HA had a MW in the 2 MDa range, indicating that it was high MW HA. Since the HA band in the gel cannot be identified using an antibody‐based method, we digested the supernatant from the stimulated fibroblasts with hyaluronidase. Upon running the gel, the band disappeared, confirming that high MW HA was produced.
5. CONCLUSION
HA is well known as an essential component of the ECM affecting both cellular function and matrix homeostasis. In addition, topical applications of HA provide important hydrative properties to the skin. Recent research has demonstrated the importance of high MW HA in this capacity—its ability to create an anti‐inflammatory environment conducive to regenerative functions contrasts with low MW HA, which is associated with a pro‐inflammatory milieu. Using a series of in vitro experiments, a formulation was developed with novel peptides and active agents that appears to stimulate the formation of high MW HA by dermal fibroblasts. This formulation design holds the promise of surface hydration using high MW HA and the intrinsic production of high MW by fibroblasts in the depths of the skin, constituting a first of its kind hydration technology. Results of multi‐center trials will be published soon.
AUTHORS CONTRIBUTIONS
ADW developed peptide, developed the science, and designed studies, analysis, and paper writing. MZ involved in data analysis, paper writing, and study design. JG involved in formulation design. DR, JV, and MP involved in paper writing contribution (clinical study to follow). MB involved in data analysis and paper writing.
ETHICAL APPROVAL
This study was undertaken as purely in vitro investigations and therefore no IRB or Ethics Committee was involved.
Widgerow AD, Ziegler ME, Garruto JA, et al. Designing topical hyaluronic acid technology—Size does matter…. J Cosmet Dermatol. 2022;21:2865–2870. doi: 10.1111/jocd.15027
Funding information
Funding applied for these studies was from Alastin, a Galderma Company
DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available from the corresponding author upon reasonable request.
REFERENCES
- 1. Rivers DA, Stern R. Hyaluronan and the process of aging in skin. In: Farage M, Miller K, Maibach H, eds. Textbook of Aging Skin. Springer; 2017. [Google Scholar]
- 2. Shatirishvili M, Burk AS, Franz CM, et al. Epidermal‐specific deletion of CD44 reveals a function in keratinocytes in response to mechanical stress. Cell Death Dis. 2016;7:e2461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Litwiniuk M, Krejner A, Grzela T. Hyaluronic acid in inflammation and tissue regeneration. Wounds. 2016;28:78‐88. [PubMed] [Google Scholar]
- 4. Rayahin JE, Buhrman JS, Zhang Y, et al. High and low molecular weight hyaluronic acid differentially influence macrophage activation. ACS Biomater Sci Eng. 2015;1:481‐493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Griss J, Viteri G, Sidiropoulos K, et al. ReactomeGSA – efficient multi‐omics comparative pathway analysis. Mol Cell Proteomics. 2020;19:2115‐2125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Saito S, Takayama Y, Mizumachi K, et al. Lactoferrin promotes hyaluronan synthesis in human dermal fibroblasts. Biotech Lett. 2011;33:33‐39. [DOI] [PubMed] [Google Scholar]
- 7. Takayama Y. Effects of lactoferrin on skin wound healing. Biochem Cell Biol. 2012;90:87‐100. [DOI] [PubMed] [Google Scholar]
- 8. Campiche R, Jackson E, Laurent G, et al. Skin filling and firming activity of a hyaluronic acid inducing synthetic tripeptide. Int J Pept Res Ther. 2019;26:181‐189. [Google Scholar]
- 9. Product monograph; Nagase Chemtex PIPS; Phosphatidylserine & phosphatidylinositol; 2015.
- 10. Product monograph sodium hyaluronate crosspolymer hylasome EG10 vantage; 2020.
- 11. Li H, Lee HS, Kim SH, et al. Antioxidant and anti‐inflammatory activities of methanol extracts of Tremella fuciformis and its major phenolic acids. J Food Sci. 2014;79:C460‐C468. [DOI] [PubMed] [Google Scholar]
- 12. Liao WC, Hsueh CY, Chan CF. Antioxidative activity, moisture retention, film formation, and viscosity stability of Auricularia fuscosuccinea, white strain water extract. Biosci Biotechnol Biochem. 2014;78:1029‐1036. [DOI] [PubMed] [Google Scholar]
- 13. Product monograph: Symdecanox, Symrise; 2015.
- 14. Kim KH, Son ED, Kim HJ, et al. EGR3 is a late epidermal differentiation regulator that establishes the skin‐specific gene network. J Invest Dermatol. 2019;139:615‐625. [DOI] [PubMed] [Google Scholar]
- 15. Jang M, Baek S, Kang G, et al. Dissolving microneedle with high molecular weight hyaluronic acid to improve skin wrinkles, dermal density and elasticity. Int J Cosmet Sci. 2020;42:302‐309. [DOI] [PubMed] [Google Scholar]
- 16. Ruppert SM, Hawn TR, Arrigoni A, et al. Tissue integrity signals communicated by high‐molecular weight hyaluronan and the resolution of inflammation. Immunol Res. 2014;58:186‐192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Widgerow AD, Moradi A, Poehler J. A double‐blind randomized controlled trial evaluating the efficacy and tolerability of a topical body treatment in combination with cryolipolysis procedures. J Drugs Dermatol. 2019;18:342‐348. [PubMed] [Google Scholar]
- 18. Moradi A, Poehler J, Bell M. A randomized double‐blind trial evaluating the efficacy and tolerability of topical body treatment with TriHex Technology® combined with abdomen cryolipolysis or radiofrequency procedures. J Cosmet Dermatol. 2020;19:677‐681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Widgerow AD, Ziegler ME, Casas LA. Topical Skin Treatment and Its Influence on Surgical Healing: Review of Literature and Underlying Physiology. Aesthet Surg J Open Forum. 2021;3:ojab029. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.