

Abstract
Chinese fir (Cunninghamia lanceolata) is one of southern China's most important native tree species, which has experienced noticeable climate-induced changes. Published papers (1978–2020) on tree growth of Chinese fir forests in China were collected and critically reviewed. After that, a comprehensive growth data set was developed from 482 sites, which are distributed between 102.19° and 130.07°E in longitude, between 21.87° and 37.24°N in latitude and between 5 and 2260 m in altitude. The dataset consists of 2265 entries, including mean DBH (cm), mean H (m), volume (m3), biomass (dry weight) (kg) (stem (over bark) biomass, branches biomass, leaves biomass, bark biomass, aboveground biomass, roots biomass, total trees biomass) and related information, i.e. geographical location (Country, province, study site, longitude, latitude, altitude, slope, and aspect), climate (mean annual precipitation-MAP and mean annual temperature-MAT), stand description (origin, age, canopy density and stand density), and sample regime (plot size, number and investigation year). Our results showed that (1) the best prediction of height was obtained using nonlinear composite model Height = , (R2 = 0.8715, p < 0.05), (2) the equation Volume = DBH2/(387.8 + 19,190/Height) (R2 = 0.9833, p < 0.05) was observed to be the most suitable model for volume estimation. Meanwhile, when the measurements of the variables are difficult to carry out, the volume model Volume = 0.03957 − 0.01215*DBH + 0.00118*DBH2 (R2 = 0.9573, p < 0.05) is determined from DBH only has a practical advantage, (3) the regression equations of component biomass against DBH explained more significant than 86% variability in almost all biomass data of woody tissues, which were ranked as total trees (97.25%) > aboveground (96.55%) > stems (with bark) (96.17%) > barks (88.95%) > roots (86.71%), and explained greater than 64% variability in branch biomass. The foliage biomass equation was the poorest among biomass components (R2 = 0.6122). The estimation equations derived in this study are particularly suitable for the Chinese fir forests in China. This dataset can provide a theoretical basis for predicting and assessing the potential of carbon sequestration and afforestation activities of Chinese fir forests on a national scale.
Subject terms: Ecology, Ecology
Introduction
Forests cover c. 1/3 of the land area1, c. 85% of global above-ground carbon (C)2. Forests have considerable potential to mitigate human-caused climate change3–6. Afforestation and forest management are the leading measures in increasing carbon sink and mitigating greenhouse gas concentrations7,8. Afforestation is also an important way to restore the ecological environment in southern China9.
Chinese fir (Cunninghamia lanceolata), one of the most important native tree species in southern China, has been widely planted because of its fast growth and high-yield10. The Chinese fir plantation area is c. 11 million hectares, accounting for c. 12.9% of the plantation forest in China11,12. The carbon storage of Chinese fir forests is 63.69 Tg accounting for 1.71% of China's forests carbon storage13.
During the past years, Chinese fir tree growth has experienced noticeable climate-induced changes14–17. Therefore, understanding growth characteristics is crucial to managing and predicting Chinese fir forests under future climate change. The growth rate of Chinese fir forests is an essential indicator for evaluating the forest restoration process and carbon dioxide storage potential.
Allometry, linking easily measurable variables such as diameter at breast height (DBH) and tree height (H) with other structural and functional characteristics of trees, is the most reliable and commonly used method for estimating forest biomass, volume, and net primary productivity, etc.18–20. There has been some synthetic research on Chinese fir in recent years. For example, the relationships: DBH, H and volume, Number of samples (N) = 182 (Location: 29°8′N, 118°24′E)21, N = 44 (Location: 27°45′N, 109°10′E)22, N = 39923 and N = 184024 (Location: 25°16′–26°46′N 107°55′–109°36′E, Guizhou Province); DBH, H and biomass (Stems, barks, branches, leaves and roots), N = 20 (Location: 26°25′–27°04′N, 117°05′–117°40′E)25, N = 39 (Location: 26°25′–27°04′N, 117°05′–117°40′E)26; DBH, H and biomass (Stems, barks, branches, leaves, roots and total trees), N = 6 (Location: 26°28′N, 117°57′E)27, N = 10 (Location: 29°05′–29°23′N, 119°10′–119°20′E)28, N = 18 (Location: 31°10′–31°20′N, 115°30′–115°50′E)29; DBH, H and biomass (Stems, barks, branches, leaves), N = 600 (Location: 20°12′–34°59′N, 97°23′–122°18′E)30. As mentioned above, almost all the biomass investments were carried out using local observations, and the national-scale biomass equation of Chinese fir was rare. In addition, no biomass equations for roots and total trees can be used for national-scale estimates. Furthermore, the existing volume equations were all local-scale volume models, while the national-scale volume model was still unknown. To implement global forest carbon sink monitoring and assessment, developing a single tree species allometric growth equation suitable for large-scale estimation has become a trend31. Here, we established a comprehensive dataset of Chinese fir growth in China (N = 2265). The sample plots cover almost all the distribution areas of Chinese fir. The data of DBH and H was from the field measurement, and the data of volume and biomass was from the felled trees. In this study, national-scale allometric models were developed to estimate the DBH-Height relationships, stem volume, and biomass (Stems (with bark) plus barks, branches, foliage, aboveground, roots and total trees) for Chinese fir trees.
Materials and methods
Literature retrieval
Published studies (1978–2020) collected from available online full-text databases, including Academic Resource Search (https://scholar.lanfanshu.cn/), Sci-Hub Literature Search (https://sci-hub.se/). China Knowledge Resource Integrated Database (http://www.cnki.net/), Wanfang Data Knowledge Service Platform (http://www.wanfangdata.com/), Baidu Academic (https://xueshu.baidu.com/), Springer Link (http://link.springer.com/), and Docin (https://www.docin.com/). Different combinations of the keywords “Chinese fir” (or “Cunninghamia lanceolata”) with “DBH”, “height”, “volume”, and “biomass” were searched. We made a great effort to compile a complete growth dataset of planted and natural Chinese fir in Asia. The “Dataset” and “References cited in the dataset” are stored in Excel xlsx format. These references were published publicly, and the data in these papers were allowed to be cited. The collection and processing of data are permitted by the laws of the People's Republic of China.
Data collection
From the literature, data was only used if all of the following criteria were met to obtain reliable growth data: Chinese fir monoculture plantations and natural forests (pure forest stand or its proportion exceeds 70%); When the study consists of multiple treatments, only data from the control treatment was selected, DBH and H were averaged from the measurement values of all trees in plots or with a random or systematic sampling method. Calculate the volume and biomass of a single tree based on the felled wood sample (allometry-derived data was excluded); The forest stands included in the dataset were limited to those that have not been disturbed by fire, pests, or recently logging; Data have undergone substantial checking, for example, a cross-check for relevant information from different sources and preliminary correlation analysis among growth variables.
Therefore, 2265 records were used to generate a comprehensive growth dataset of Chinese fir. The data set includes mean DBH (cm), mean H (m), volume (m3), biomass (kg) (dry weight) (stem (over bark) biomass, branches biomass, leaves biomass, bark biomass, aboveground biomass, roots biomass, total biomass). In addition, it also contains the following related information, geographical location (Country, province, study site, longitude, latitude, altitude, slope, and aspect), when geographic coordinates were not available in the original papers, Google Earth (Version: 7.3.4.8428) was applied to find longitude, latitude and altitude. Climate (mean annual precipitation-MAP and mean annual temperature-MAT), stand description (origin, age, canopy density and stand density), and sample regime (plot size, number and investigation year). These variables and their definitions, units, number, and range are listed in Table 1.
Table 1.
Variable information in the data set.
| Column code | Definition | Unit | Number | Range |
|---|---|---|---|---|
| ID | Unique identification number of each record | N/A | 2265 | 1–2265 |
| Province | Province location of study site | N/A | 15 | N/A |
| Study site | Locality name of study site | N/A | 482 | N/A |
| Latitude | Latitude of study site | ° | 2265 | 21.87°–37.24°N |
| Longitude | Longitude of study site | ° | 2265 | 102.19°–130.07° E |
| Altitude | Altitude of study site | m | 2257 | 5–2260 |
| Aspect | Slope direction of study site, including none (flat slope), sunny, half-sunny, shady, half-shady | N/A | 804 | N/A |
| Slope | Slope degree | ° | 1120 | 0–62 |
| Origin | Natural or planted forests | N/A | 2265 | N/A |
| MAT | Mean annual temperature | °C | 2265 | 11.9–28 |
| MAP | Mean annual precipitation | mm | 2265 | 837–2900 |
| Age | Stand age | years | 1507 | 1–180 |
| Height | Mean tree height | m | 2252 | 0.58–36.4 |
| DBH | Mean diameter at breast height | cm | 2265 | 0.9–69.2 |
| Vtree | Mean tree volume (felled wood) | m3/tree | 423 (1260 trees) | 0.00039–2.0915 |
| Vstand | Stand volume (felled wood data) | m3 ha−1 | 309 | 1.77–1185 |
| Stems | Stems (over bark) biomass (dry weight, felled wood) | kg | 406 (1287 trees) | 0.01–466.05 |
| Bark | Bark biomass (dry weight, felled wood) | kg | 204 (438 trees) | 0.00–57.83 |
| Branches | Branches biomass (dry weight, felled wood) | kg | 359 (1215 trees) | 0.02–43.56 |
| Leaves | Leaves biomass (dry weight, felled wood) | kg | 378 (1234 trees) | 0.07–19.58 |
| Aboveground | Aboveground biomass (dry weight, felled wood) | kg | 356 (1260 trees) | 0.15–497.88 |
| Root | Root biomass (dry weight, felled wood) | kg | 338 (1030 trees) | 0.03–79.98 |
| Total | Total tree biomass (dry weight, felled wood) | kg | 347 (1179 trees) | 0.17–577.86 |
| Density | Stand density | trees ha−1 | 1732 | 15–10,000 |
| Area | Plot area | m2 | 1508 | 40–669,000 |
| Plot | Plot numbers, i.e., replications | N/A | 1506 | 1–342 |
| Year | Investigation year | N/A | 1278 | 1978–2020 |
| Reference | Data sources | N/A | 688 | N/A |
Site and climate
The data was derived from 482 research sites in China (102.19°–130.07° E; 21.87°–37.24°N), including Anhui, Fujian, Guangdong, Guangxi, Guizhou, Henan, Hubei, Hunan, Jiangsu, Jiangxi, Shandong, Sichuan, Taiwan, Yunnan, Zhejiang provinces (Fig. 1). The average annual precipitation (MAP) ranges from 837 to 2900 mm, and the average annual temperature (MAT) ranges from 11.9 to 29 °C.
Figure 1.
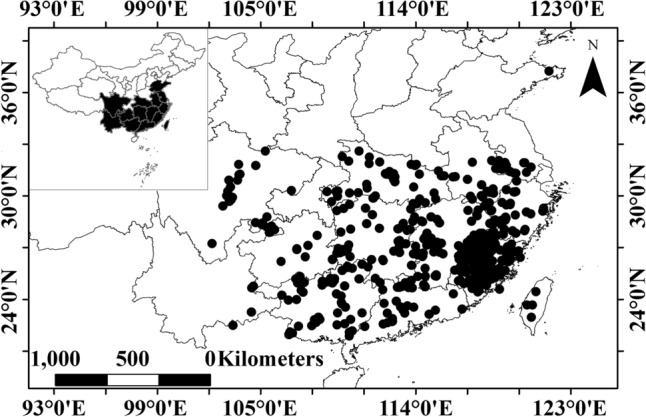
Study sites of Chinese fir in China, including Anhui, Fujian, Guangdong, Guangxi, Guizhou, Henan, Hubei, Hunan, Jiangsu, Jiangxi, Shandong, Sichuan, Taiwan, Yunnan, Zejiang provinces. The x-axis is longitude and y-axis is latitude. This figure was generated by the software QGIS 3.10.14 (https://www.qgis.org/en/site/).
Data estimates and evaluation
A total of 2252 available pairs of mean DBH and mean tree height in the data set were used to establish the DBH-H correlation with a power function (), R2 = 0.8715, P < 0.05, see Fig. 2 and Table 2). To calculate tree volume from only one known variable of DBH, tree height was firstly calculated with the power H–DBH equation in Fig. 2. The Matlab software version 2021a was used for this and subsequent analyses.
Figure 2.
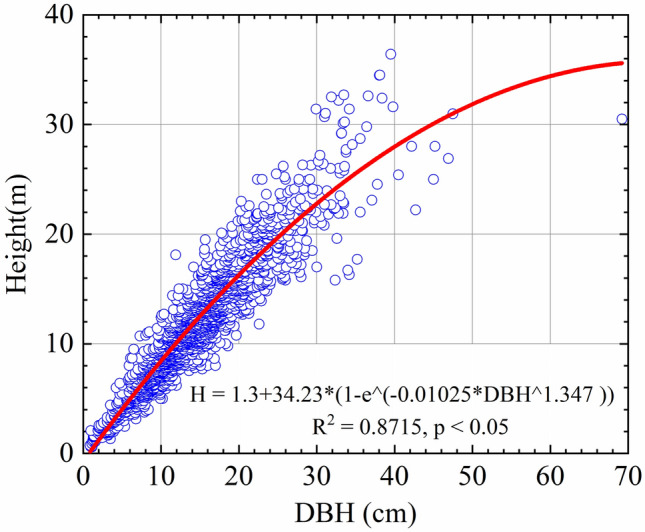
Relationship between diameter at breast height (DBH) and mean tree height (H) in the data set, Height = (R2 = 0.8715, p < 0.05).
Table 2.
Coefficient and fit statistics of different DBH-height equations.
| Model no. | Equation | a | b | c | R2 | RMSE |
|---|---|---|---|---|---|---|
| 1 | H = 1.3 + a*(DBHb) | 0.7738 | 0.98 | 0.8585 | 2.074 | |
| 2 | H = 1.3 + (a*DBH/(b + DBH)) | 293.1 | 378.5 | 0.8607 | 2.057 | |
| 3 | H = 1.3 + a*(1 − e(−b*DBH)) | 145.1 | 0.005311 | 0.8609 | 2.056 | |
| 4 | H = 1.3 + (DBH2/(a + b*DBH)2) | 2.462 | 0.1341 | 0.8703 | 1.985 | |
| 5 | H = 1.3 + a*e(b/DBH) | 36.02 | − 16.94 | 0.8518 | 2.122 | |
| 6 | H = 1.3 + (a*DBH /(DBH + 1)) + b*DBH | − 0.2143 | 0.7403 | 0.8583 | 2.075 | |
| 7 | H = 1.3 + a*((DBH /(1 + DBH))b) | 37.22 | 18.07 | 0.8551 | 2.098 | |
| 8 | H = 1.3 + a*(1 − e(−b*DBHc)) | 34.23 | 0.01025 | 1.347 | 0.8715 | 1.977 |
| 9 | H = 1.3 + (DBH 2/(a + b*DBH + c*DBH2)) | 4.967 | 0.7804 | 0.01505 | 0.8705 | 1.983 |
| 10 | H = 1.3 + a*DBH*(e(−b*DBH)) | 0.7714 | 0.002671 | 0.861 | 2.055 | |
| 11 | H = 1.3 + a*DBH + b*DBH2 | 0.7735 | − 0.0021 | 0.8612 | 2.053 | |
| 12 | H = 1.3 + a*e((b/DBH) +c) | 2.223 | − 7.68 | 2.234 | 0.683 | 3.103 |
| 13 | H = 1.3 + a*e(−(b/DBHc)) | 177 | 7.819 | 0.3849 | 0.8699 | 1.988 |
The volume equations were classified as two types by the independent variables: volume = f(DBH) and volume = f(DBH, height). These equations have been previously applied for tree volume models in forestry32–35. There were three equations in f(DBH) and four equations in f(DBH, height), and the parameter estimates and fit statistics were computed by all equations (Table 3). Equation (7) was evaluated as the most suitable volume equation with the highest coefficient of determination (R2) and lowest root mean square error (RMSE) (Fig. 3). The estimated stand volume was determined by multiplying the optimal estimated tree volume by the stand density.
Table 3.
Coefficient and fit statistics of different stem volume equations.
| Model no | Equation | a | b | c | d | R2 | RMSE | DBH range (cm) |
|---|---|---|---|---|---|---|---|---|
| 1 | V = a + bDBH2 | − 0.06187 | 0.00087 | 0.9489 | 0.05274 | 3.6–47.5 | ||
| 2 | V = a + bDBH + cDBH2 | 0.03957 | − 0.01215 | 0.00118 | 0.9573 | 0.04822 | 3.6–47.5 | |
| 3 | V = aDBH + bDBH2 | − 0.00783 | 0.001075 | 0.9562 | 0.04882 | 3.6–47.5 | ||
| 4 | V = a + bDBH2H | 0.02093 | 3.37e−05 | 0.9788 | 0.03399 | 3.6–47.5 | ||
| 5 | V = aDBH2H | 3.513e−05 | 0.9733 | 0.03808 | 3.6–47.5 | |||
| 6 | V = a + bDBHcHd | 79.22 | − 79.4 | − 0.00082 | − 0.00106 | 0.3421 | 0.1892 | 3.6–47.5 |
| 7 | V = DBH2/(a + b/H) | 387.8 | 19,190 | 0.9833 | 0.03017 | 3.6–47.5 |
Figure 3.
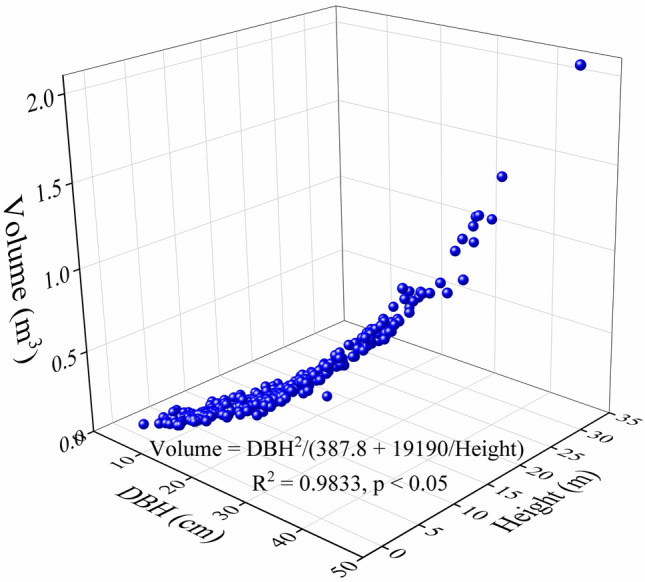
Relationships of Volume against diameter at breast height (DBH) and Height, Volume = DBH2/(387.8 + 19,190/Height) (R2 = 0.9833, p < 0.05).
There is no universally accepted equation form for biomass prediction. Most biomass equations in scientific literature adopt the power function of M = aDb36–39, and this biomass model was also used in this paper. The regression equations of component biomass against DBH explained more significant than 86% variability in almost all biomass data of woody tissues, including total trees, aboveground, roots, stems (with bark), barks, and explained greater than 64% variability in branches biomass (Table 4; Fig. 4). The foliage biomass equation was the poorest among biomass components (R2 = 0.6122) (Table 4; Fig. 4).
Table 4.
Allometric equations relating biomass components (kg) to diameter at breast height (DBH, cm).
| Component | a | b | R2 | RMSE | DBH range (cm) |
|---|---|---|---|---|---|
| Stems (with bark) | 0.02106 | 2.8 | 0.9617 | 12.91 | 1.31–36.41 |
| Barks | 0.006084 | 2.566 | 0.8895 | 3.556 | 4.8–36.41 |
| Branches | 0.04026 | 1.9 | 0.6481 | 4.57 | 1.19–27.3 |
| Leaves | 0.1681 | 1.233 | 0.6122 | 1.913 | 1.19–27.3 |
| Aboveground | 0.04305 | 2.624 | 0.9655 | 13.87 | 1.31–36.41 |
| Roots | 0.02184 | 2.311 | 0.8671 | 5.259 | 1.31–36.41 |
| Total trees | 0.06986 | 2.53 | 0.9725 | 15.29 | 1.19–36.41 |
Figure 4.

Relationships of component biomass against diameter at breast height. (a–g) Stand for stem (with bark), total branch, total foliage, aboveground, total root, and total biomass, respectively.
Discussion
DBH–height relationship is widely used to estimate timber volume, biomass, and other important parameters for forest growth and yield in forest management40. Accurate tree height prediction is critical in forest modeling, inventory and management decision making41. Our results show that DBH and tree height are well correlated at the national scale, and the DBH-height model displayed a good fit (R2 = 0.8715, p < 0.05). Therefore, such national-scale relationships will avoid much cost of constructing site-specific relations42. Furthermore, it is especially useful to improve stand volume and carbon stock estimation in national-scale forest inventory.
The R2 value (R2 = 0.9833, p < 0.05) for our national-scale two-variable volume equation was similar to other local scale volume equations reported by Li21 (R2 = 0.9820), Zeng et al.23 (R2 = 0.9994), and Xia et al.24 (R2 = 0.9868). Models that incorporate DBH and height usually give good-fits43–46. Although the best-fit model to estimate stem wood volume was a two-variable volume equation (Table 3), in some cases, this two-variable model is not practical because the measurements of these variables are difficult to carry out with high accuracy, particularly in closed forests47. Besides, the results of this study verified that a one-variable equation with DBH only can also be used to get good estimates of Chinese fir tree volume in China (R2 = 0.9573, p < 0.05). Meanwhile, the DBH is easy to measure accurately in the field, so this one-variable equation has a practical advantage.
We developed a set of biomass equations for Chinese fir trees growing in China. In the previous studies, Li and Zhao30 used 600 Chinese fir trees for aboveground biomass equations, while the biomass dataset used in our study consisted of 1525 Chinese fir trees. A larger sample size can reduce parameter estimation uncertainty48. Compared with the previous equations30 developed for Chinese fir forests on a national scale, the new system of equations included two more biomass components-roots and total trees (Table 4). Belowground biomass comprises about 25% of total biomass in forest ecosystems49. However, because of the laborious and time-consuming50, direct measurements of belowground biomass are seldom51. Precise quantification of underground carbon storage in forest ecosystems is of great significance for effectively predicting how future environmental changes will affect global carbon dynamics52. There is an urgent need to develop some algorithm for estimating this carbon pool. Correlations found in this study indicate that equations using dbh as the predictive variable can offer a good estimate of the stems, total aboveground, roots, and total trees biomass, but a poor estimate of branches and foliage biomass (Table 4), which is consistent with other findings that a single DBH-based allometric equation provides a reasonable prediction for total aboveground53,54, root55, and total tree biomass56. The improved predictability of total and aboveground biomass may also be attributable to the fact that most biomass exists within the stem component that is, in itself, highly correlated with dbh. Poor predictions for branches and foliage biomass may result from variation in biomass allocation due to soil conditions57, stand age58 and stand density59, which is consistent with the previous studies57,60,61.
Conclusions
In this study, 13 DBH-Height models were tested on trees in Chinese-fir forests between 1 and 180 years old in southern China. Model selection was based on goodness of fit. The best prediction of height was obtained using nonlinear composite model H = , R2 = 0.8715, P < 0.05, which used three parameters, this was recommended for Chinese-fir forests in China.
We also used seven models to test suitability for Chinese fir tree volume estimation. Of these, the equation Volume = DBH2/(387.8 + 19,190/Height) (R2 = 0.9833, p < 0.05) was observed to be the most suitable model for volume estimation. In addition, when the measurements of the variables are difficult to carry out, the volume model (Volume = 0.03957 − 0.01215*DBH + 0.00118*DBH2) is determined from DBH only has a practical advantage.
In addition, the regression equations of component biomass against DBH explained more significant than 86% variability in almost all biomass data of woody tissues, which were ranked as total trees (97.25%) > aboveground (96.55%) > stems (with bark) (96.17%) > barks (88.95%) > roots (86.71%), and explained greater than 64% variability in branch biomass. The foliage biomass equation was the poorest among biomass components (R2 = 0.6122).
As mentioned above, these estimation equations derived in this study are particularly suitable for the Chinese fir forests in China.
Supplementary Information
Acknowledgements
We would like to acknowledge many scientists and researchers whose field measurements made this workpossible. This research was supported by the National Natural Science Foundation of China (No. 42177454).
Author contributions
Y.J., Z.H.,J.Z.,and S.H. conceived the idea; Y.J. download the papers; Y.J., Z.H. and Z.H. analyzed data; Y.J. wrote the manuscript; Z.H. and L.H. writing—review and editing; J.Z. and Z.H. reviewed and approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Data availability
The datasets used and/or analysed during the current study are available from the corresponding author upon reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Yangao Jiang, Email: jiangyangao-jyg@163.com.
Zhiguang Han, Email: hanzg@henu.edu.cn.
Junhui Zhang, Email: jhzhang@iae.ac.cn.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-022-22809-6.
References
- 1.FAO . The State of the World’s Forests 2018—Forest Pathways to Sustainable Development. FAO; 2018. [Google Scholar]
- 2.Pan Y, et al. A large and persistent carbon sink in the world’s forests. Science. 2011;333(6045):988–993. doi: 10.1126/science.1201609. [DOI] [PubMed] [Google Scholar]
- 3.Luyssaert S, et al. Tradeoffs in using European forests to meet climate objectives. Nature. 2018;562(7726):259–262. doi: 10.1038/s41586-018-0577-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Büntgen U, et al. Limited capacity of tree growth to mitigate the global greenhouse effect under predicted warming. Nat. Commun. 2019 doi: 10.1038/s41467-019-10174-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Anderegg WRL, et al. Climate-driven risks to the climate mitigation potential of forests. Science. 2020;368:1327. doi: 10.1126/science.aaz7005. [DOI] [PubMed] [Google Scholar]
- 6.Jiang M, et al. The fate of carbon in a mature forest under carbon dioxide enrichment. Nature. 2020;580(7802):227–231. doi: 10.1038/s41586-020-2128-9. [DOI] [PubMed] [Google Scholar]
- 7.Naudts K, et al. Europe’s forest management did not mitigate climate warming. Science. 2016;351(6273):597–599. doi: 10.1126/science.aad7270. [DOI] [PubMed] [Google Scholar]
- 8.Tong X, et al. Forest management in southern China generates short term extensive carbon sequestration. Nat. Commun. 2020 doi: 10.1038/s41467-019-13798-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yu K, Yao X, Deng Y, Lai Z, Lin L, Liu J. Effects of stand age on soil respiration in Pinus massoniana plantations in the hilly red soil region of Southern China. CATENA. 2019;178:313–321. doi: 10.1016/j.catena.2019.03.038. [DOI] [Google Scholar]
- 10.Mei G, Sun Y, Sajjad S. Models for predicting the biomass of Cunninghamia lanceolata trees and stands in southeastern China. PLoS ONE. 2017;12:e0169747. doi: 10.1371/journal.pone.0169747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wu H, et al. Soil phosphorus bioavailability and recycling increasedwith stand age in Chinese fir plantations. Ecosystems. 2019;23:973–988. doi: 10.1007/s10021-019-00450-1. [DOI] [Google Scholar]
- 12.State Forestry Administration. General situation of forest resources in China. The 8th National Forest Inventory (State Forestry Administration, 2014).
- 13.Wang X, et al. Vegetation carbon storage and density of forest ecosystems in China. Chin. J. Appl. Ecol. 2001;12(1):13–16. [PubMed] [Google Scholar]
- 14.Kang H, et al. Simulating the impact of climate change on the growth of Chinese fir plantations in Fujian province, China. NZ J. For. Sci. 2017;47(1):20. doi: 10.1186/s40490-017-0102-6. [DOI] [Google Scholar]
- 15.Lu Y, et al. A process-based approach to estimate Chinese fir (Cunninghamia lanceolata) distribution and productivity in southern China under climate change. Forests. 2015;6:360–379. doi: 10.3390/f6020360. [DOI] [Google Scholar]
- 16.Zhang X, et al. Relative contributions of competition, stand structure, age, and climate factors to tree mortality of Chinese fir plantations: Long-term spacing trials in southern China. For. Ecol. Manag. 2020;465:118103. doi: 10.1016/j.foreco.2020.118103. [DOI] [Google Scholar]
- 17.You R, et al. Variation in wood physical properties and effects of climate for different geographic sources of Chinese fir in subtropical area of China. Sci. Rep. 2021 doi: 10.1038/s41598-021-83500-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Djomo AN, Ibrahima A, Saborowski J, Gravenhorst G. Allometric equations for biomass estimations in Cameroon and pan moist tropical equations including biomass data from Africa. For. Ecol. Manag. 2010;260(10):1873–1885. doi: 10.1016/j.foreco.2010.08.034. [DOI] [Google Scholar]
- 19.Peng D, et al. Estimating the aboveground biomass for planted forests based on stand age and environmental variables. Remote Sens. 2019;11(19):2270. doi: 10.3390/rs11192270. [DOI] [Google Scholar]
- 20.Zhou X, Yang M, Liu Z, Li P, Xie B, Peng C. Dynamic allometric scaling of tree biomass and size. Nat. Plants. 2021;7(1):42–49. doi: 10.1038/s41477-020-00815-8. [DOI] [PubMed] [Google Scholar]
- 21.Li, L. Study on the tree volume table compilation of Chinese fir in Kaihua Forest Farm (Beijing Forestry University, 2011) http://cdmd.cnki.com.cn/Article/CDMD-10022-1011134655.htm (in Chinese).
- 22.Wang JP, et al. Study on the effect of Chinese fir volume formula on estimating the volume of fir standing trees in different sites. Guizhou For. Technol. 1991;19(1):26–29. [Google Scholar]
- 23.Zeng WS, et al. Compatible tree volume and aboveground biomass equations for Chinese fir plantation in Guizhou. J. Beijing For. Univ. 2011;33(4):1–6. [Google Scholar]
- 24.Xia ZS, et al. Construction of tree volume equations for Chinese fir plantation in Guizhou Province, southwestern China. J. Beijing For. Univ. 2012;34(1):1–5. [Google Scholar]
- 25.Lin H. Study on biomass and carbon storage of main coniferous forest in Jiangle state-owned forestry farm. J. Fujian For. Sci. Technol. 2018;45(1):30–34. doi: 10.13428/j.cnki.fjlk.2018.01.007. [DOI] [Google Scholar]
- 26.Cai, Z. A study on biomass models of Cunninghamia lanceolata plantation in Fujian. (Beijing Forestry University, 2014), http://cdmd.cnki.com.cn/Article/CDMD-10022-1014327550.htm (in Chinese).
- 27.Chen G, et al. Carbon storage in a chronosequence of Chinese fir plantations in southern China. For. Ecol. Manag. 2013;300:68–76. doi: 10.1016/j.foreco.2012.07.046. [DOI] [Google Scholar]
- 28.Zhang G, et al. Biomass Characteristics of dominant tree species (group) at Lingnan forest farm in Anhui province. Scientia Silvae Sinicae. 2012;48(5):136–140. doi: 10.1007/s11783-011-0280-z. [DOI] [Google Scholar]
- 29.Shi W, et al. Biomass model and carbon storage of Chinese fir plantation in Dabieshan Mountains in Anhui. Resour. Environ. Yangtze Basin. 2015;24(5):758–764. doi: 10.11870/cjlyzyyhj201505007. [DOI] [Google Scholar]
- 30.Li H, Zhao P. Improving the accuracy of tree-level aboveground biomass equations with height classification at a large regional scale. For. Ecol. Manag. 2013;289:153–163. doi: 10.1016/j.foreco.2012.10.002. [DOI] [Google Scholar]
- 31.Zeng W, Tang S. A new general allometric biomass model. Nat. Precedings. 2011 doi: 10.1038/npre.2011.6704.1. [DOI] [Google Scholar]
- 32.Schumacher FX, Hall FDS. Logarithmic expression of timber-tree volume. J. Agric. Res. 1933;47(9):719–734. [Google Scholar]
- 33.Honer TG. A new total cubic foot volume function. For. Chron. 1965;41(4):476–493. doi: 10.5558/tfc41476-4. [DOI] [Google Scholar]
- 34.Burkhart HE. Cubic-foot volume of loblolly pine to any merchantable top limit. South. J. Appl. For. 1977;2:7–9. doi: 10.1093/sjaf/1.2.7. [DOI] [Google Scholar]
- 35.Lee D, Seo Y, Choi J. Estimation and validation of stem volume equations for Pinus densiflora, Pinus koraiensis, and Larix kaempferi in South Korea. For. Sci. Technol. 2017;13(2):77–82. doi: 10.1080/21580103.2017.1315963. [DOI] [Google Scholar]
- 36.Chen BH, Chen CY. A preliminary study on the biomass and productivity of Picea koraiensis forests in the dunes. Scientia Silvae Sinicae. 1980;4:269–278. [Google Scholar]
- 37.Niklas KJ. Plant Allometry: The Scaling of Form and Process. University of Chicago Press; 1994. [Google Scholar]
- 38.Ketterings QM, et al. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. For. Ecol. Manag. 2001;146:199–209. doi: 10.1016/S0378-1127(00)00460-6. [DOI] [Google Scholar]
- 39.Chen XG. The biomass and allometric equation of a 20-years-old Cunninghamia lanceolata plantation. Prot. For. Sci. Technol. 2007;4:28–29, 40. doi: 10.3969/j.issn.1005-5215.2007.04.010.(inChinese). [DOI] [Google Scholar]
- 40.Wang XP, et al. Climatic control of primary forest structure and DBH–height allometry in Northeast China. For. Ecol. Manag. 2006;234:264–274. doi: 10.1016/j.foreco.2006.07.007. [DOI] [Google Scholar]
- 41.Peng C, et al. Developing and evaluating tree height–diameter models at three geographic scales for black spruce in Ontario. N. J. Appl. For. 2004;21(2):83–92. doi: 10.1093/njaf/21.2.83. [DOI] [Google Scholar]
- 42.López-Serrano FR, et al. Site and weather effects in allometries: A simple approach to climate change effect on pines. For. Ecol. Manag. 2005;215(1–3):251–270. doi: 10.1016/j.foreco.2005.05.014. [DOI] [Google Scholar]
- 43.Zhang C, et al. Developing aboveground biomass equations both compatible with tree volume equations and additive systems for single trees in Poplar plantations in Jiangsu Province, China. Forests. 2016;7:32. doi: 10.3390/f7020032. [DOI] [Google Scholar]
- 44.Liu JC, et al. Comparing non-destructive methods to estimate volume of three tree taxa in Beijing, China. Forests. 2019;10:92. doi: 10.3390/f10020092. [DOI] [Google Scholar]
- 45.Thangjam U, et al. Developing tree volume equation for Parkia timoriana grown in home gardens and shifting cultivation areas of North-East India. For. Trees Livelihoods. 2019;28(12):1–13. doi: 10.1080/14728028.2019.1624200. [DOI] [Google Scholar]
- 46.Dutcă I, et al. Does slope aspect affect the aboveground tree shape and volume allometry of European Beech (Fagus sylvatica L.) trees? Forests. 2022;13:1071. doi: 10.3390/f13071071. [DOI] [Google Scholar]
- 47.Segura M, Kanninen M. Allometric models for tree volume and total aboveground biomass in a tropical humid forest in Costa Rica. Biotropica. 2005;37(1):2–8. doi: 10.2307/30045500. [DOI] [Google Scholar]
- 48.Wang XW, et al. Additive tree biomass equations for Betula platyphylla Suk. plantations in Northeast China. Ann. For. Sci. 2018;75:60. doi: 10.1007/s13595-018-0738-2. [DOI] [Google Scholar]
- 49.Niklas KJ, Enquist BJ. Canonical rules for plant organ biomass partitioning and annual allocation. Am. J. Bot. 2002;89(5):812–819. doi: 10.3732/ajb.89.5.812. [DOI] [PubMed] [Google Scholar]
- 50.Xiang WH, et al. General allometric equations and biomass allocation of Pinus massoniana trees on a regional scale in southern China. Ecol. Res. 2011;26:697–711. doi: 10.1007/s11284-011-0829-0. [DOI] [Google Scholar]
- 51.Brown S. Measuring carbon in forests: Current status and future challenges. Environ. Pollut. 2002;116:363–372. doi: 10.1016/s0269-7491(01)00212-3. [DOI] [PubMed] [Google Scholar]
- 52.Brassard BW, et al. Influence of environmental variability on root dynamics in northern forests. Crit. Rev. Plant Sci. 2009;28:179–197. doi: 10.1080/07352680902776572. [DOI] [Google Scholar]
- 53.Montagu KD, et al. Developing general allometric relationship for regional estimates of carbon sequestration—An example using Eucalyptus pilularis from seven contrasting sites. For. Ecol. Manag. 2005;204:113–127. doi: 10.1016/j.foreco.2004.09.003. [DOI] [Google Scholar]
- 54.Williams RJ, et al. Allometry for estimating aboveground tree biomass in tropical and subtropical eucalypt woodlands: Towards general predictive equations. Aust. J. Bot. 2005;53:607–619. doi: 10.1071/BT04149. [DOI] [Google Scholar]
- 55.Ouimet R, et al. Estimation of coarse root biomass and nutrient content for sugar maple, jack pine, and black spruce using stem diameter at breast height. Can. J. For. Res. 2008;38:92–100. doi: 10.1139/x07-134. [DOI] [Google Scholar]
- 56.Peichl M, Arain MA. Allometry and partitioning of above-and belowground tree biomass in an age-sequence of white pine forests. For. Ecol. Manag. 2007;253:68–80. doi: 10.1016/j.foreco.2007.07.003. [DOI] [Google Scholar]
- 57.Bond-Lamberty B, et al. Aboveground and below-ground biomass and sapwood area allometric equations for six boreal tree species of northern Manitoba. Can. J. For. Res. 2002;32:1441–1450. doi: 10.1139/x02-063. [DOI] [Google Scholar]
- 58.King JS, et al. Biomass partitioning in red pine (Pinus resinosa) along a chronosequence in the Upper Peninsula of Michigan. Can. J. For. Res. 2007;37(1):93–102. doi: 10.1139/x06-217. [DOI] [Google Scholar]
- 59.Ziania D, Mencuccini M. Aboveground biomass relation-ships for beech (Fagus moesiaca Cz.) trees in Vermio Mountain, northern Greece, and generalised equations for Fagus sp. Ann. For. Sci. 2003;60(5):439–448. doi: 10.1051/forest:2003036. [DOI] [Google Scholar]
- 60.Martin JG, et al. Aboveground biomass and nitrogen allocation of ten deciduous southern Appalachian tree species. Can. J. For. Res. 1998;28(11):1648–1659. doi: 10.1139/x98-146. [DOI] [Google Scholar]
- 61.Wang C. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For. Ecol. Manag. 2006;222:9–16. doi: 10.1016/j.foreco.2005.10.074. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author upon reasonable request.