
Figure 1. Differential effects of PG01042, PG01037, and VK4-116 on G protein-dependent signaling in HEK-293T cells transfected with D3R and D1R.
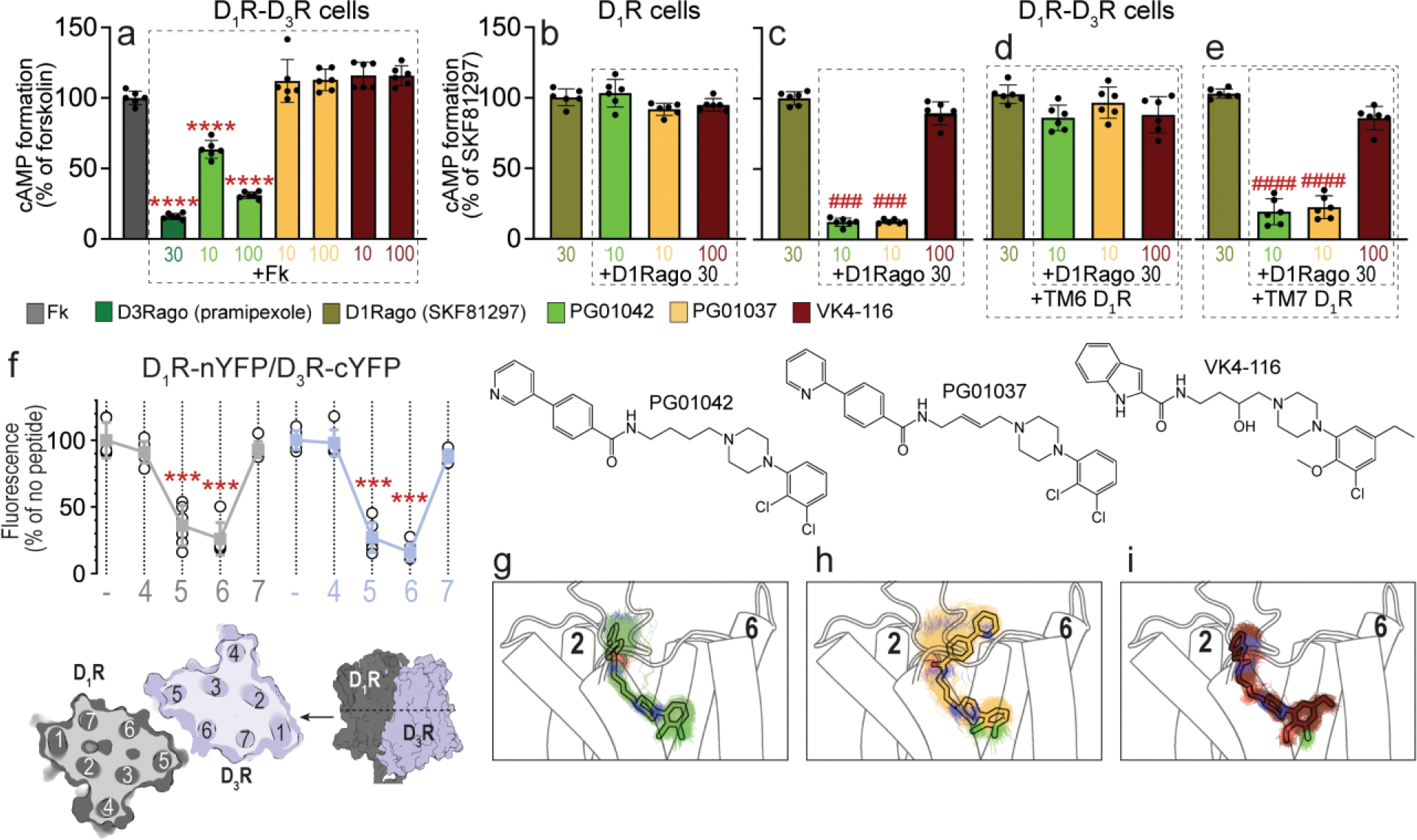
a-e. Results from cAMP formation experiments in HEK-293T cells transfected with D1R-Rluc cDNA (1 μg) with or without D3R-YFP cDNA (1.5 μg) (D1R-D3R cells and D1R cells, respectively). In a, cells are treated with the D2-like receptor agonist pramipexole (D3Rago; 30 nM for 10 min) or the D3R ligands PG01042, PG01037 and VK4-116 (all at 10 or 100 nM for 15 min) before forskolin (Fk, 0.5 μM). In b-e, cells are pre-treated or not with D1R TM6 or TM7 peptides (4 μM for 4 h) and treated with PG01042 (10 nM), PG01037 (10 nM) and VK4-116 (100 nM) for 15 min before the D1R agonist SKF81297 (30 nM; D1Rago). Values of cAMP formation are shown as mean ± S.D. (n = 6) and expressed as percentage of Fk-treated or D1Rago-treated cells in each condition (100% represents 80–100 pmols cAMP/106 cells). ****: p < 0.0001 versus FK; ### and ####: p < 0.001 and p < 0.0001 versus D1Rago, respectively (one-way ANOVA followed by Tukey’s post hoc comparisons). f. Results from BiFC experiments in HEK-293T cells co-transfected with D1R-nYFP and D3R-cYFP in the absence (−) or the presence of the indicated TM peptides (at 4 μM) from D1R (gray symbols and plots) or D3R (blue symbols and plots). Fluorescence values (in means ± S.D.) are expressed as the percentage of the fluorescence in the absence (−) of the indicated TM peptides (n = 6, with triplicates); ***: p < 0.001 versus control values (one-way ANOVA followed by Dunnett’s post hoc comparisons). The schemes illustrate extracellular and parallel to the membrane views of the computational model of the D1R-D3R heteromer built using the TM 5/6 interface (see text). g-i. Representative structures (solid sticks) and evolution (lines) of PG01042 (g, in green), PG01037 (h, in orange) and VK4-116 (i, in purple) in complex with D3R (white cylinders, only the initial structure is shown) as devised from three replicas of unbiased 1μs MD simulations. The second pharmacophore unit of PG01042 and VK4-116 remained stable at the ECD near TM 2, whereas this part of PG01037 favors its interaction with TM 6 (see Suppl. Fig. 2).